Embed Size (px)
Citation preview

Vasoactivity of Intraluminal and Extraluminal Agonists inPerfused Retinal Arteries
Dao-Yi Yu, Valerie A. Alder, Stephen J. Cringle, Er-Ning Su, and Paula K. Yu
Purpose. To evaluate the vasoactive response of isolated perfused arteries of the pig to K+
and adrenergic agonists and to compare the effects of intraluminal (IL) and extraluminal(EL) drug delivery.
Methods. A new microperfusion system was developed, in which short lengths of porcineretinal arteries (outer diameter 90.4 ± 2.7 fim) were cannulated at both ends and perfusedat a controlled rate (5 /xl/min) with outflow through a single side branch. The diameter ofthe vessel and the intraluminal pressure were monitored, and the effect of intraluminally andextraluminally applied agonists was determined. Endothelial cell function and the integrityof the blood retinal barrier was verified.
Results. Consistent vasoactive responses were obtained from most vessels. The resting diameterof the vessel was not greatly influenced by changes in flow rate or intraluminal pressure overthe physiological range. Adrenaline and noradrenaline caused dose-dependent contractions,which were larger when applied intraluminally than they were when applied extraluminally.The largest contraction for adrenaline was 19.0% ± 2.1% (n = 13) IL and 8.4% ± 1.5% (n= 13) EL, and for noradrenaline, 17.8% ± 1.9% (n = 13) IL and 6.8% ± 1.1% (n = 13) EL.The IL contraction to 124-mM K+, 19.0% ± 1.6% (n = 21), was also greater than that forEL application, 5.0% ± 1.0% (n = 13). We found that the existence of myogenic contractionswas restricted to the special case in which vessels with no branches were pressurized underzero flow conditions.
Conclusions. Pig retinal arteries exhibited asymmetry in their responses to adrenergic agonistsand K+, with contractions significantly larger when the drug was applied to the intraluminalsurface rather than the extraluminal surface. This asymmetry may reflect an important prop-erty of retinal vessels. Microperfusion systems of this type may prove valuable in developinga better understanding of control mechanisms in retinal circulations. Invest Ophthalmol VisSci. 1994; 35:4087-4099.
A he retinal circulation is a highly specialized vascularbed where the requirement of guaranteeing a goodoptical image appears to have taken precedence overthe provision of a plentiful blood supply. It is relativelysparse, and even under normal conditions it operateswith a large arteriovenous oxygen difference. Themain arteries divide to form two to three capillary bedsin most holangiotic mammalian retinas. Adrenergic
From the Lions Eye Institute, University of Western Australia, Nedlands, Australia.Supported by the Juvenile Diabetes Foundation International, the Medical ResearchFund of Western Australia, and the National Health and Medical Research Councilof Australia.Submitted for publication October 19, 1993; revised June 7, 1994; accepted June10, 1994.Proprietary interest category: N.Reprint requests: Dr. Dao-Yi Yu, Lions Eye Institute, University of WesternAustralia, Nedlands, WA 6009, Australia.
autonomic innervation ceases at the optic nerve head,1
and there is no evidence of parasympathetic or peptid-ergic innervation in retinal arteries,2 so the dynamicprocess of matching local tissue blood flow to localmetabolic needs must be controlled by other factors.Likely candidates for such control mechanisms are:smooth muscle myogenic responses to blood pressureand intraocular pressure changes, release of metabolicsignalers from neural and glial tissue, blood-borne fac-tors, autocoids such as nitric oxide and endothelin,which may be released from endothelial cells, andfeedback communication between the venous and ar-terial sides of the circulation. It is possible that consid-erable heterogeneity exists in the vasoactive responseof different segments of the retinal circulation fromfirst order arteriole to capillary and retinal veins. Loca-
Investigative Ophthalmology & Visual Science, November 1994, Vol. 35, No. 12Copyright © Association for Research in Vision and Ophthalmology 4087

4088 Investigative Ophthalmology & Visual Science, November 1994, Vol. 35, No. 12
tion-specific heterogeneity has already been reportedin in vitro investigations of the ophthalmociliary ar-tery,3 and the observation of a differential shunting ofthe red cell moiety of blood between the superficialand deep capillary beds in the retina4 also implies lo-calized control mechanisms. If such heterogeneity isa consistent feature of the retinal circulation, then invitro preparations could provide the means to helpunravel this differential control. The local control ofretinal arteries has already been explored in in vitropreparations of the larger (>200 //m diameter) bo-vine retinal arteries, using a ring segment technique.5"12 The ring segment technique that we and othershave used has the limitation that candidate vasoactiveagents act on both sides of the vessel wall simultane-ously, and that small diameter vessels such as thosefound in the human cannot be used. The recent pro-liferation of papers demonstrating differential re-sponses to intraluminal (IL) and extraluminal (EL)application of vasoactive candidates in vessels fromother organs such as the brain,1314 ear,!5 tail,16 mesen-tery,17 and cheek pouch,1819 as well as the carotid ar-tery,20 has driven us to question whether retinal vesselsdisplay a preference for IL or EL application of candi-date vasoactive agents. To differentiate between IL-and EL-induced responses, it is necessary to perfusethe vessel segment with a physiological solution so thatthe agents can be added either to the perfusate or tothe bathing solution separately. This constant perfu-sion technique is arguably closer to the true physiolog-ical situation than can be achieved with the ring seg-ment preparation or with vessels that are pressurizedunder no flow conditions.21 Indeed, perfusion may befunctionally important to vasoactivity, becausechanges in flow through vessels have been shown tomediate vessel responses both in vivo and in vitro.22
We have therefore performed pilot studies thathave allowed us to develop a perfusion-based tech-nique for retinal vessels based on the work of Chonkoet al23 in the kidney, and Duling and Rivers24 for mes-enteric vessels. It also owes some of its features to themicroperfusion system used by Murta et al25 for bloodretinal barrier studies in rabbit retinal vessels. Al-though this study relates exclusively to retinal vesselsfrom pigs, the same system has proved suitable forretinal vessels from human, cat, and dog. We initiallyconcentrated on optimizing and validating the perfu-sion technique itself and then investigated the differ-ential sensitivity of in vitro pig retinal arteries to IL andEL application of catecholamines and K+. Histology ofthese vessels has been performed in parallel so that thestructure of the vessel wall may be related to vascularresponses. We regard this as a necessary first step inthe process of determining controlling factors for theretinal circulation.
MATERIALS AND METHODS
Animals
A total of 60 pig eyes was used in this study. All proce-dures adhered to the ARVO Statement for the Use ofAnimals in Ophthalmic and Vision Research. The eyeswere obtained either from a local abattoir immediatelyafter slaughter or from animals under Halothane anes-thesia in our own surgical facility. In both instancesthe eyes were enucleated and placed in a sealed bottleof oxygenated Krebs solution containing 0.6% dia-lyzed albumin (BSA) and kept on ice for transfer tothe laboratory (10 to 45 minutes).
Histology of the Retinal Artery
In a preliminary histologic study, two freshly enucleatedpig eyes were used for structural studies of the retinalartery. The ophthalmic artery was cannulated, and thewhole eye was perfused initially with Krebs solution for10 minutes at an input pressure of 50 to 70 mm Hg,followed by 2.5% gluteraldehyde for 30 minutes. Aftermaking an entry hole into the vitreous at pars planaciliaris to allow easy diffusion into the ocular tissues, thewhole eyeball was further fixed in 2.5% gluteraldehydefor 24 hours. Pieces of major arteries and adjacent retinawere excised, post fixed in osmium tetroxide, dehy-drated in graded ethanol's, and embedded in epoxyresin for light and electron microscopy.
Preparation of Vessel for In Vitro Study
Using a dissecting microscope (Zeiss, Oberkochen, Ger-many), the eyes were sectioned at pars plana ciliaris,separating the anterior segment and adherent vitreousbody from the posterior pole. The retina, choroid, andsclera were divided into quadrants using a razor blade,taking care not to section any major retinal arteries.The retina was carefully separated from the underlyingchoroid and sclera using an iris speculum. A quadrantof retina was placed on a hollowed glass slide containingKrebs solution plus 0.6% dialyzed albumin (BSA) keptat less than 4°C. Using a combination of transmittedlight with a stereo microscope (Wild, M3Z, Heerbrugg,Switzerland), individual first-order retinal arteries (out-side diameter 60 to 120 //m) were dissected from retinaltissue with a fire-polished micropipette. A segment ofartery approximately 800 to 1500 //m long was selected;in most experiments, care was taken to include a sidebranch of outside diameter 15 to 30 fim, although in afew cases, a segment with no side branch was chosen.This arterial segment was then transferred to the incuba-tor chamber.
Incubator Chamber
The incubator chamber (PDMI-2, Medical Systems,New York, NY) was mounted on the stage of an in-verted microscope (Nikon Diaphot-TMD, Tokyo, Ja-
Vasoactivity of Perfused Pig Retinal Arteries 4089
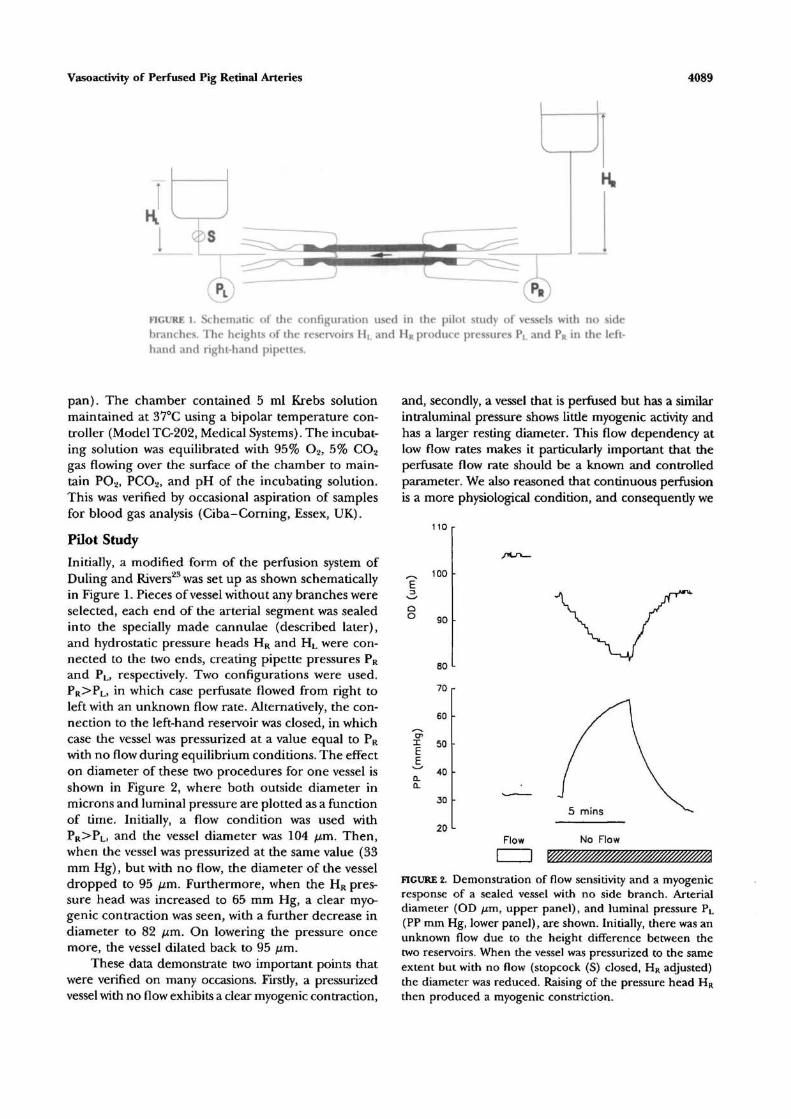
FIGURE l. Schematic of the configuration used in the pilot study of vessels with no sidebranches. The heights of the reservoirs HL and HR produce pressures PL and PR in the left-hand and right-hand pipettes.
pan). The chamber contained 5 ml Krebs solutionmaintained at 37°C using a bipolar temperature con-troller (Model TC-202, Medical Systems). The incubat-ing solution was equilibrated with 95% O2, 5% CO2
gas flowing over the surface of the chamber to main-tain PO2, PCO2, and pH of the incubating solution.This was verified by occasional aspiration of samplesfor blood gas analysis (Ciba-Corning, Essex, UK).
Pilot Study
Initially, a modified form of the perfusion system ofDuling and Rivers23 was set up as shown schematicallyin Figure 1. Pieces of vessel without any branches wereselected, each end of the arterial segment was sealedinto the specially made cannulae (described later),and hydrostatic pressure heads HR and HL were con-nected to the two ends, creating pipette pressures PR
and PL, respectively. Two configurations were used.PR>PL, in which case perfusate flowed from right toleft with an unknown flow rate. Alternatively, the con-nection to the left-hand reservoir was closed, in whichcase the vessel was pressurized at a value equal to PR
with no flow during equilibrium conditions. The effecton diameter of these two procedures for one vessel isshown in Figure 2, where both outside diameter inmicrons and luminal pressure are plotted as a functionof time. Initially, a flow condition was used withPR>PL, and the vessel diameter was 104 fj,m. Then,when the vessel was pressurized at the same value (33mm Hg), but with no flow, the diameter of the vesseldropped to 95 fim. Furthermore, when the HR pres-sure head was increased to 65 mm Hg, a clear myo-genic contraction was seen, with a further decrease indiameter to 82 jum. On lowering the pressure oncemore, the vessel dilated back to 95 /im.
These data demonstrate two important points thatwere verified on many occasions. Firstly, a pressurizedvessel with no flow exhibits a clear myogenic contraction,
and, secondly, a vessel that is perfused but has a similarintraluminal pressure shows little myogenic activity andhas a larger resting diameter. This flow dependency atlow flow rates makes it particularly important that theperfusate flow rate should be a known and controlledparameter. We also reasoned that continuous perfusionis a more physiological condition, and consequently we
110
100
90
80
70
60
40
30
20Flow No Flow
FIGURE 2. Demonstration of flow sensitivity and a myogenicresponse of a sealed vessel with no side branch. Arterialdiameter (OD fxm, upper panel), and luminal pressure PL
(PP mm Hg, lower panel), are shown. Initially, there was anunknown flow due to the height difference between thetwo reservoirs. When the vessel was pressurized to the sameextent but with no flow (stopcock (S) closed, HR adjusted)the diameter was reduced. Raising of the pressure head HR
then produced a myogenic constriction.

4090 Investigative Ophthalmology & Visual Science, November 1994, Vol. 35, No. 12
FIGURE 3. Schematic of the cannulation and perfusion system for the isolated retinal arteries.Both ends of the vessel were cannulated and sealed by gentle squeezing the vessel wallbetween the inner and outer pipettes. Syringe pumps delivered perfusate at a controlledrate. Pressure transducers PI, P2, P3, and P4 measured the resulting pressures at the pointsindicated. Typically, the right-hand pipette system was used for the main perfusate (5 jj,\/min) and drug delivery, whereas the left-hand pipette was used to measure the intravascularpressure. The only perfusate outflow path (arrow) is through the side branch of the mainvessel. Test agents were introduced into the perfusate stream through a sample injectorvalve, VI. The vessel was maintained in an incubation bath on the stage of an invertedmicroscope (M), and video images were output to a monitor and automated vessel diametermeasuring system. All relevant parameters were recorded on a chart recorder, and datawere streamed directly to computer disk when required.
devised a perfusion system in which the flow could becontrolled and the pressures and the vessel diametercontinuously monitored. Because this is a new tech-nique, we describe it in some detail.
Cannula and Perfusion System
General. The principle used in these experimentswas that vessels with a side branch were cannulated atboth ends and perfused through one end (proximal)with a flow of 5 fxl/min in the orthograde direction,with the side branch acting as the exit route, as shownin Figure 3. This choice of 5 fi\/min as a baselineperfusion flow rate was based on in vivo measurementsusing laser Doppler velocimetry.26 The other end (dis-tal) was generally left with a small flow rate (0.3 yul/min). This residual flow helped ensure that drugs de-livered from the other pipette did not collect in thedistal end of the vessel. The vessel diameter to theright-hand side of the branch was monitored continu-ously. The diameter changes in response to intralumi-nal or extraluminal delivery of drugs were compared.
Cannula. Each cannula consisted of four concen-tric pipettes (Fig. 3). The outer or holding pipette Awas shaped on its innermost surface to present twoconstrictions. The constriction closer to the open endof the pipette served as a surface against which theinner pipette B, the pipette that entered the vessellumen, squeezed and sealed the vessel wall. The inner-most constriction served to centralize the tip of pipette
B and to separate it from the inner wall of A. PipetteD delivered the perfusate and IL drugs directly at theshoulder of the perfusing pipette B, close to the vesselto minimize the delay between injection and arrivalof the drug in the vessel. Pipette C also delivered per-fusate at a rate one fifth of that in D. This ensuredthat there was no back flow of drugs into the mainbody of B that could later serve as a pocket of contami-nation. The resistance of pipette B was minimized andoffered less resistance to perfusate flow than that pre-sented by a typical second-order branch.
Perfusion and Mounting System, Perfusate flow wasdelivered by a computer-controlled syringe pump(Model 22, Harvard Apparatus, South Natick, MA) con-taining one 1-ml and one 5-ml gas-tight syringe (Activon;Pennant Hills, New South Wales, Australia) connectedto die C and D pipettes, respectively. A specially manu-factured pipette holding system was developed that al-lowed relative movement between pipettes A and B whilemaintaining a pressure seal and minimal compliance.The whole assembly was mounted on a joystick-con-trolled KYZ micro drive (Fine Science Tools, Foster City,CA) and angled at 35° to the horizontal. An equivalentsystem of pipettes and manipulators was used for theleft-hand side and perfused by a second, independentlycontrolled pump.
Vessel CannulationThe vessel was positioned horizontally in the incuba-tion bath close to the bottom of the dish. By conven-

Vasoactivity of Perfused Pig Retinal Arteries 4091
tion, the proximal end of the vessel was mounted onthe right-hand pipette system. The XYZ drive was ma-nipulated to place the tip of pipette A adjacent tothe proximal end of the vessel. Relative movementbetween pipettes A and B was achieved using a hydrau-lic micro drive. Pipette B was retracted into the can-nula and gentle suction was applied to pipette A todraw the end of the vessel into the cavity between thetwo constrictions. Pipette B was then advanced intothe vessel lumen such that it squeezed the vessel wallagainst the constriction. This procedure was per-formed while a continuous perfusion of 5 //1/min wasflowing. It was found that this helped keep the vessellumen open. Once this end of the vessel was sealed,the perfusate flow flushed out the red blood cells fromthe lumen of the vessel, and a similar procedure wasrepeated on the other end. Once both ends were can-nulated, perfusion delivery at the distal end was re-duced to a residual level, typically 0.3 //1/min. Thevessel was left to stabilize for 30 minutes.
Experiment Control and Diameter andPressure Measurement
All of the data recording and much of the instrumentcontrol was under computer control. The software wasdeveloped using the graphical programming languageLabView (National Instruments, Austin, TX) and runon a 486DX PC with the appropriate serial and IEEEcommunications cards added. The inverted micro-scope image of the vessel was captured with a CCDcamera (MW-BL602, Panasonic, Tokyo, Japan), wasdisplayed on a color video monitor (Trinitron; Sony,Osaka, Japan), and could be recorded using a video-cassette recorder (VO-9850P, Sony; Tokyo, Japan).Vessel diameter was measured by a second computer(386SX) using a commercially produced softwarepackage and frame grabbing card (DIAMTRAK, Din-dima; Ringwood, Victoria, Australia), which producedan analog output proportional to vessel diameter. Thepressures PI, P2, P3, and P4 (Fig. 3) were measured byconventional transducers (Cobe, Arvada, Colorado),each connected to a bridge amplifier (5B38-02, Ana-log Devices; Norwood, MA). The monitoring of pres-sure at two points in each pipette system was foundto be a valuable aid in detecting small leaks or partialblockages in the system. These pressure signals, to-gether with vessel diameter, the flow rate of the twopumps, and injection signals from the sample injectorvalves (V! and V2, 7725i Rheodyne; Cotati, California)were all recorded on a chart recorder (LR8100, Yoko-gawa; Tokyo, Japan). Using IEEE-GPIB communica-tion between the host computer and the chart re-corder, all eight channels were sampled every 2 sec-onds, and the data were reproduced on the computerscreen and streamed directly to a spreadsheet file asrequired.
Intraluminal and Extraluminal Drug Delivery
Drugs to be delivered intraluminally were adminis-tered as a 5-//1 bolus into the perfusate stream throughthe sample injector valves. The intrinsic design of thisinjector system permitted inclusion of the drug boluswithout any pressure artifact or problems with air bub-bles. A built-in switch allowed a signal to be generatedto indicate loading and injection phases of the proce-dure, and this signal was recorded by the chart re-corder and the computer. At a typical perfusion rateof 5 //1/min, the drug arrived at the vessel approxi-mately 90 seconds after injection. This was determinedby visual observation of dye boli. Spreading of theoriginal bolus in transit results in a slight dilution ofthe drug, but this is small compared to the log unitincrements in concentration. The size, and hence theduration of the bolus, was sufficient for the vasoactiveresponse to stabilize. Extraluminal drug delivery wasachieved by direct pipetting into the incubating solu-tion to achieve the required concentration. The bath-ing solution was flushed frequently.
Blood Retinal Barrier
The retinal circulation normally has a tight blood reti-nal barrier. Sodium fluorescein (0.05%) was injectedintraluminally in the manner described above. Usingthe epifluorescence attachment (TMD-EF, Nikon),the sodium fluorescein could be seen arriving at thevessel, and we looked carefully for leakage throughthe vessel wall and for the presence of fluoresceinwithin or between the mural cells of the vessel.
Solutions
Vessels were usually bathed and perfused with normalKrebs solution with the following composition: NaCl119 mM, KC1 4.6 mM, CaCl2 1.5 mM, MgCl2 1.2 mM,NaHCO3 15 mM, NaH2PO4 1.2 mM, Glucose 6 mM.High potassium solution (124 mM K+) was producedby equimolar substitution of Na+ with K+. Solutionswere equilibrated with 95% O2, 5% CO2.
Agonists
Agonists, (+/-)-noradrenaline HC1 (NA) and (-)-adrenaline (A) were obtained from Sigma Chemicals(St. Louis, MO) and dissolved in 0.9% NaCl. Stocksolutions of all drugs were stored at — 70°C, and freshdilutions were made daily.
Experimental Protocol
After an equilibration time of 30 minutes, three re-peated IL injections of 124-mM K+ were performedto test for vessel viability and stability. After successfulcompletion of this stage, frequently, but not always, a10~8-M Ach dilatation response was checked to ensureviability of the endothelial cells. Then IL dose re-

4092 Investigative Ophthalmology 8c Visual Science, November 1994, Vol. 35, No. 12
sponse curves were measured for the appropriate ago-nist. For each IL injection, the valve loop was loadedwith the 5 fi\ of the appropriate concentration. The5-/il bolus was switched into the perfusion line, passingthrough the pipette into the artery. As soon as theresponse stabilized, replacement of the incubating so-lution was begun to ensure that drug concentrationsin the bath remained low. After at least 10 minutes,the time required for the vessel to return to its preacti-vated state, the process was repeated with a higherconcentration of intraluminal drug. Dose concentra-tion was usually increased in log units from 10~10 to10~3 M. Before and after each type of agonist, thevalve was flushed and loaded with Krebs solution, anda control IL injection was performed. A positive con-trol response meant that further flushing of the valvewas required. Then, cumulative EL dose responsecurves were measured by adding a stepwise increasingdose to the incubating solution, whereas the vessel wasperfused IL by Krebs solution. On some occasions, asa direct confirmation of IL and EL agonist sensitivities,the agonist was first applied EL and left in the bath,and the same dose was perfused intraluminally; com-parisons of diameter changes on the proximal anddistal sides of the branch vessel were made (see later).Any vessel in which the diameter did not return towithin 10% of baseline after exposure to an agonistwas rejected.
RESULTS
Histology and Visualization of Perfused Vessel
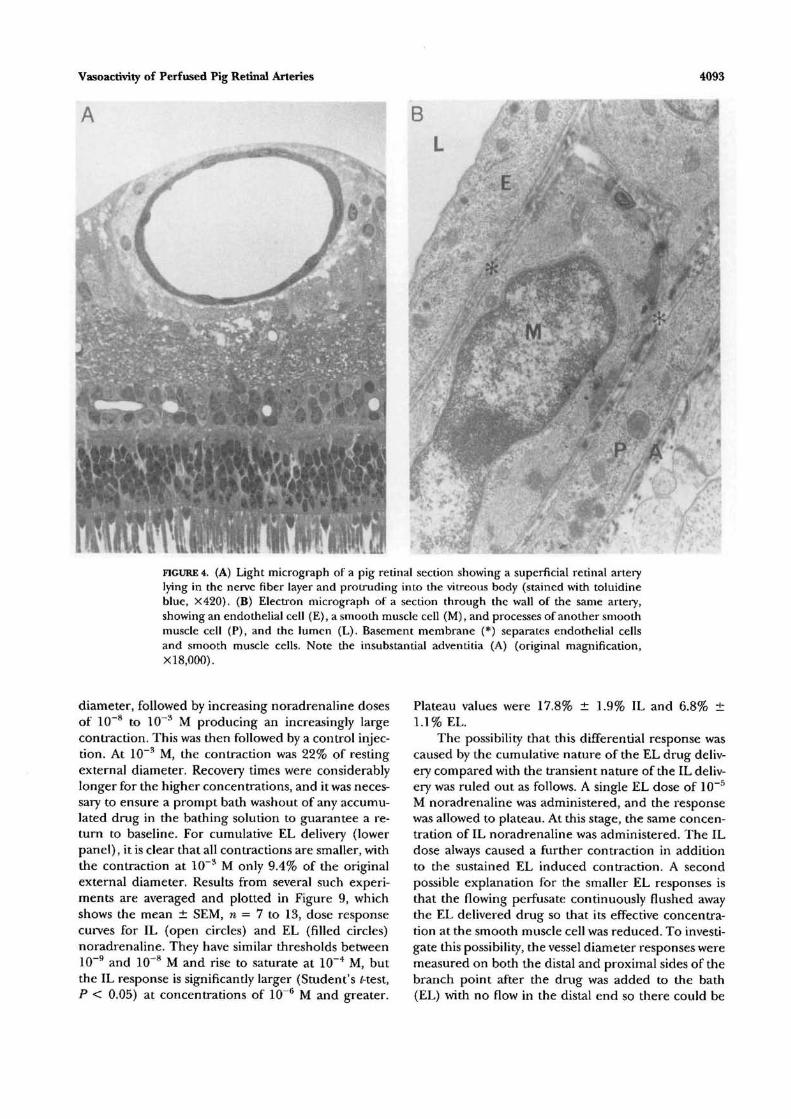
Figure 4A is a light micrograph of a pig retinal sectioncontaining a retinal artery. Perfusion fixation was per-formed at close to in vivo pressure. The retinal artery(4 to 5 mm peripheral to the disk edge) lies close tothe internal limiting membrane and protrudes intothe vitreous body; endothelial cells and smooth mus-cle cells are clearly visible. Figure 4B is an electronmicrograph of a small section of the same arterial wall.The endothelial cell (E) is supported by a peripherallylocated basement membrane (*). External to thisbasement membrane is one smooth muscle cell of themedia (SM) and its nucleus. Also present is the musclecell process (P) of another smooth muscle cell. Othersimilar sections showed that the arterial media consistsof no more than two cell bodies and a few cell pro-cesses at any location. Each smooth muscle cell is sur-rounded not only by its own plasma membrane butalso by a basement membrane that often connectswith the endothelial basement membrane of similarstructure and width. External to the smooth musclecells are loosely arranged collagen fibers that makeup the meager adventitia (A).
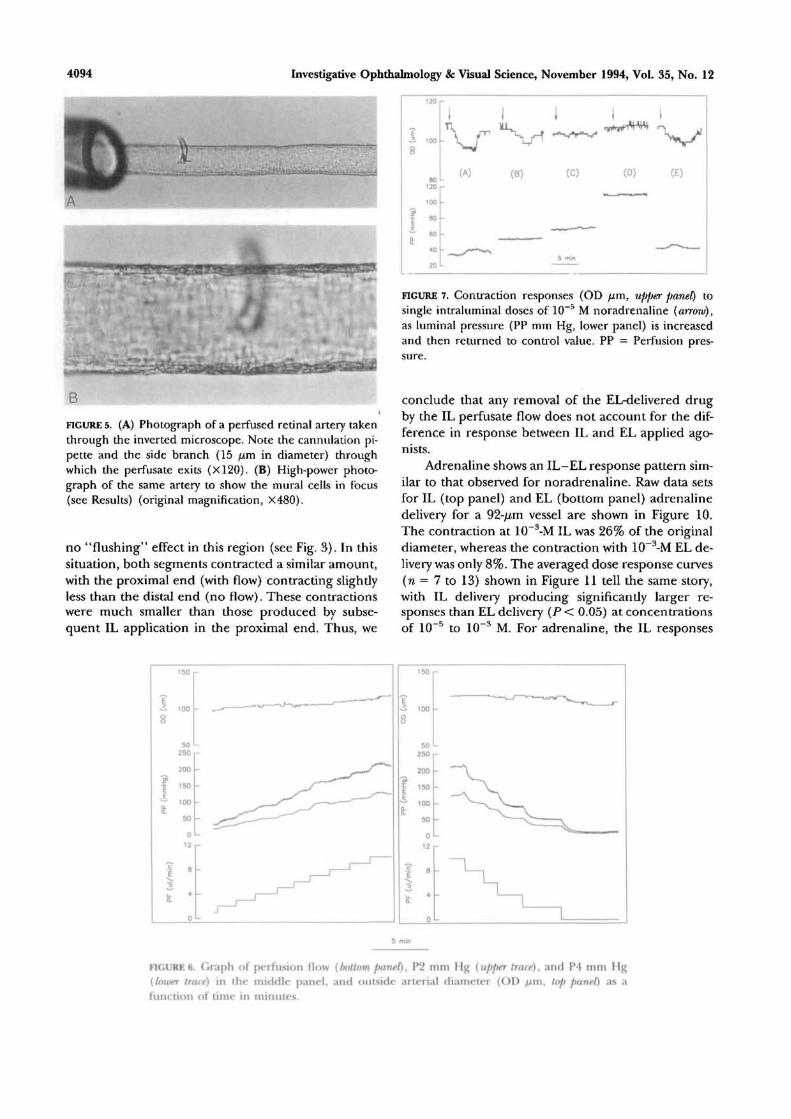
Figure 5A is a photograph of a perfused retinal
artery in situ in the incubation chamber, showing thecannulation pipette at the proximal end. The arteryhas an external diameter of 83.3 fim and a side branchof diameter 16.7 //m. A magnified photograph of thesame vessel in a different focal plane is shown in Fig-ure 5B. Note the thin muscle (media) endothelial celllayer, and the clear outer edge free of neural and glialtissue. With still higher magnification, it is possible todistinguish the intact endothelial and smooth musclecells by changing the focal plane of the inverted micro-scope. The average diameter of arteries used in thisstudy was 90.4 ± 2.7 /im (n = 42).
Effects of Perfusion Pressure and Flow on theVessel
The effect of increasing and then decreasing flow andhence intravascular pressure over a wide range is dem-onstrated in the left and right panels of Figure 6,respectively, for a vessel with a small-diameter sidebranch, so that lumen pressures are relatively high. Inthe bottom panel are the stepwise increases in flowfrom 1 //1/min to 10 /xl/min. The two traces in themiddle panel are the pressure values P2 (upper) andP4 (lower). P4 represents the IL pressure at the distalend of the vessel. P4 increases stepwise with flow. Ves-sel outside diameter (OD, top panel) increases withpressure and flow from 97 fim to 112 fim. With step-wise reduction in flow, diameter and pressure returnto similar values. The diameter of the main vessel isnot greatly influenced by changes in intraluminalpressure or flow over what one might expect to be thenormal physiological range.
However, luminal flow and pressure were im-portant parameters in determining the magnitude ofdrug responses. Figure 7 shows outside diameterchanges in response to repeated injections of 10~5 Mnoradrenaline (arrow) IL (upper panel) as luminalpressure/flow was increased (lower panel). The vesselconstricts from 108 fim to 95 fim at a pressure of 35mm Hg (A), with an accompanying small increase inpressure caused by an increase in branch resistance.For increasing values of pressure and flow, the nor-adrenaline response decreases (B) and is then lostaltogether between 65 (C) and 110 (D) mm Hg. Re-turning to lower values of pressure and flow allowsthe vessel to constrict once more in response to nor-adrenaline (E). If the luminal pressure was left highfor extended periods, vasoactivity did not recover on
. return to normal pressures.
Drug Responses
A raw dose response sequence for IL and EL deliveryof noradrenaline is presented in Figure 8 for a vesselof external resting diameter 105 fim. The top panelshows the diameter response for IL delivery. Thereis a control injection causing no change in arterial
Vasoactivity of Perfused Pig Retinal Arteries
A
4093
B
imwmmwwFIGURE 4. (A) Light micrograph of a pig retinal section showing a superficial retinal arterylying in the nerve fiber layer and protruding into the vitreous body (stained with toiuidineblue, X420). (B) Electron micrograph of a section through the wall of the same artery,showing an endothelial cell (E), a smooth muscle cell (M), and processes of another smoothmuscle cell (P), and the lumen (L). Basement membrane (*) separates endothelial cellsand smooth muscle cells. Note the insubstantial adventitia (A) (original magnification,XI 8,000).
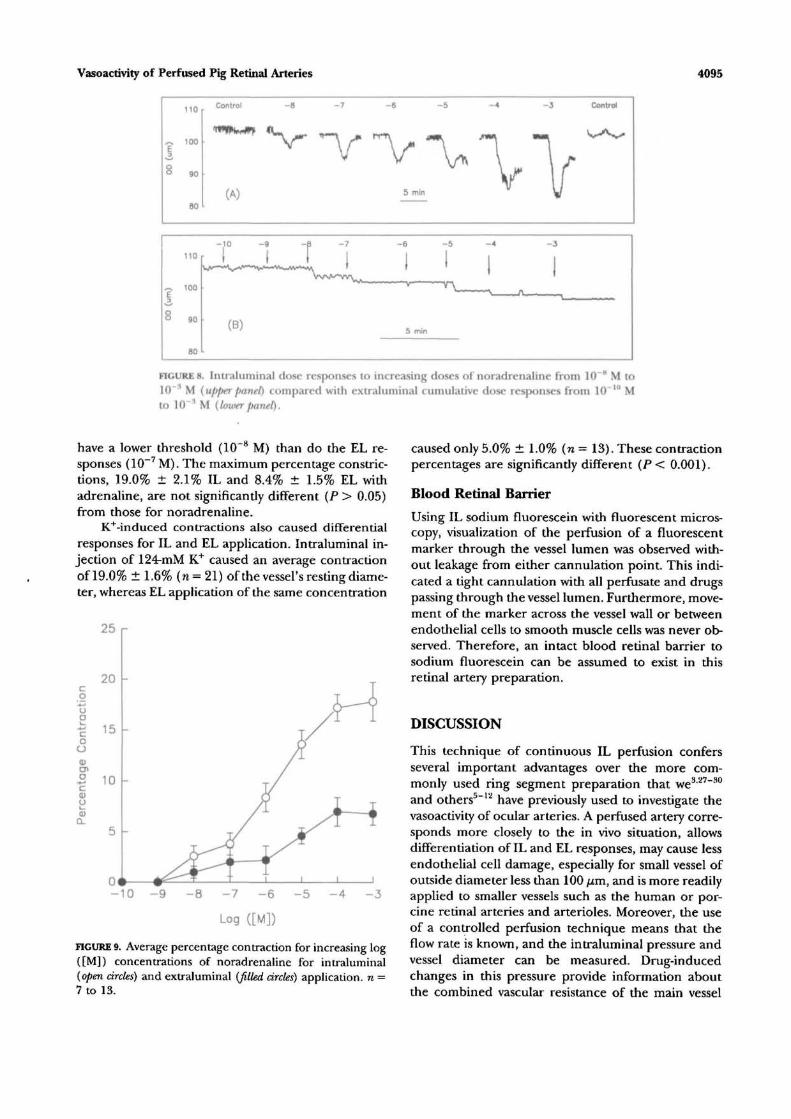
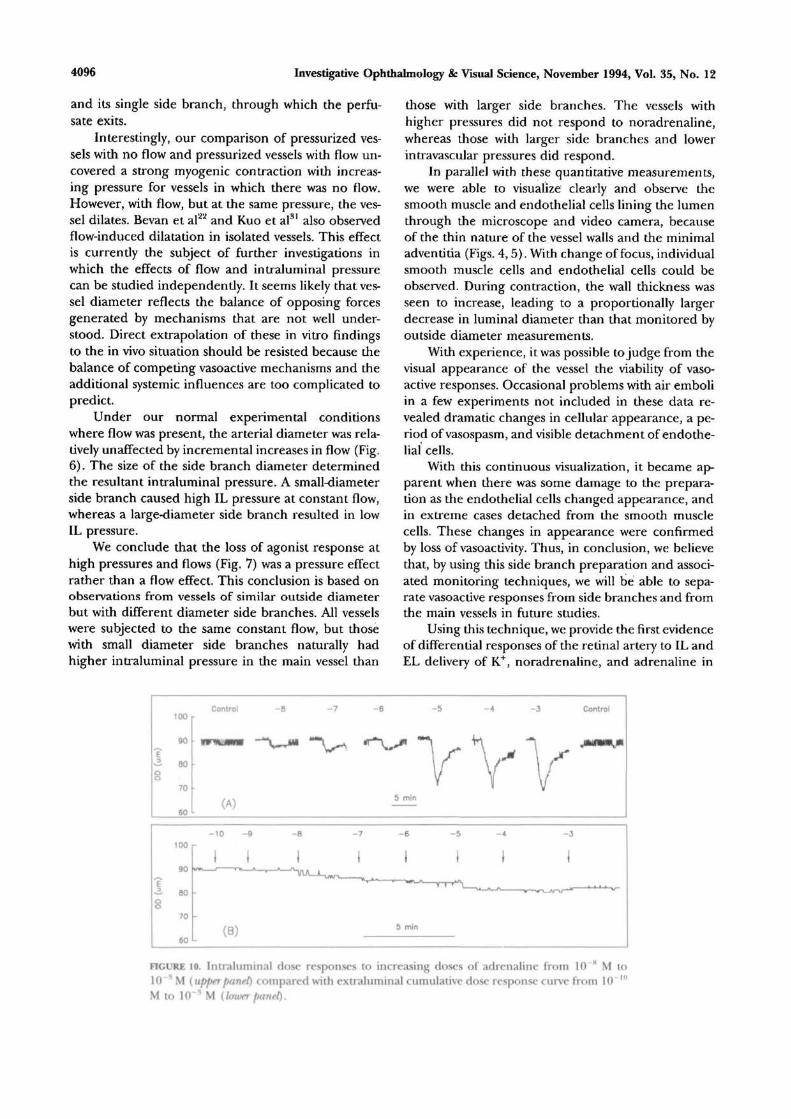
diameter, followed by increasing noradrenaline dosesof 10~8 to 1CT3 M producing an increasingly largecontraction. This was then followed by a control injec-tion. At 10~3 M, the contraction was 22% of restingexternal diameter. Recovery times were considerablylonger for the higher concentrations, and it was neces-sary to ensure a prompt bath washout of any accumu-lated drug in the bathing solution to guarantee a re-turn to baseline. For cumulative EL delivery (lowerpanel), it is clear that all contractions are smaller, withthe contraction at 10~3 M only 9.4% of the originalexternal diameter. Results from several such experi-ments are averaged and plotted in Figure 9, whichshows the mean ± SEM, n = 7 to 13, dose responsecurves for IL (open circles) and EL (filled circles)noradrenaline. They have similar thresholds between10"9 and 1(T8 M and rise to saturate at 10~4 M, butthe IL response is significantly larger (Student's Hest,P < 0.05) at concentrations of 10~6 M and greater.
Plateau values were 17.8% ± 1.9% IL and 6.8% ±1.1% EL.
The possibility that this differential response wascaused by the cumulative nature of the EL drug deliv-ery compared with the transient nature of the IL deliv-ery was ruled out as follows. A single EL dose of 10~5
M noradrenaline was administered, and the responsewas allowed to plateau. At this stage, the same concen-tration of IL noradrenaline was administered. The ILdose always caused a further contraction in additionto the sustained EL induced contraction. A secondpossible explanation for the smaller EL responses isthat the flowing perfusate continuously flushed awaythe EL delivered drug so that its effective concentra-tion at the smooth muscle cell was reduced. To investi-gate this possibility, the vessel diameter responses weremeasured on both the distal and proximal sides of thebranch point after the drug was added to the bath(EL) with no flow in the distal end so there could be
4094 Investigative Ophthalmology & Visual Science, November 1994, Vol. 35, No. 12
B
FIGURE 5. (A) Photograph of a perfused retinal artery takenthrough the inverted microscope. Note the cannulation pi-pette and the side branch (15 fim in diameter) throughwhich the perfusate exits (X120). (B) High-power photo-graph of the same artery to show the mural cells in focus(see Results) (original magnification, X480).
no "flushing" effect in this region (see Fig. 3). In thissituation, both segments contracted a similar amount,with the proximal end (with flow) contracting slightlyless than the distal end (no flow). These contractionswere much smaller than those produced by subse-quent IL application in the proximal end. Thus, we
(A) (B) (C) (D) (E)
FIGURE 7. Contraction responses (OD p , upf>er panel) tosingle intraluminal doses of 10~5 M noradrenaline {arrow),as luminal pressure (PP mm Hg, lower panel) is increasedand then returned to control value. PP = Perfusion pres-sure.
conclude that any removal of the EL-delivered drugby the IL perfusate flow does not account for the dif-ference in response between IL and EL applied ago-nists.
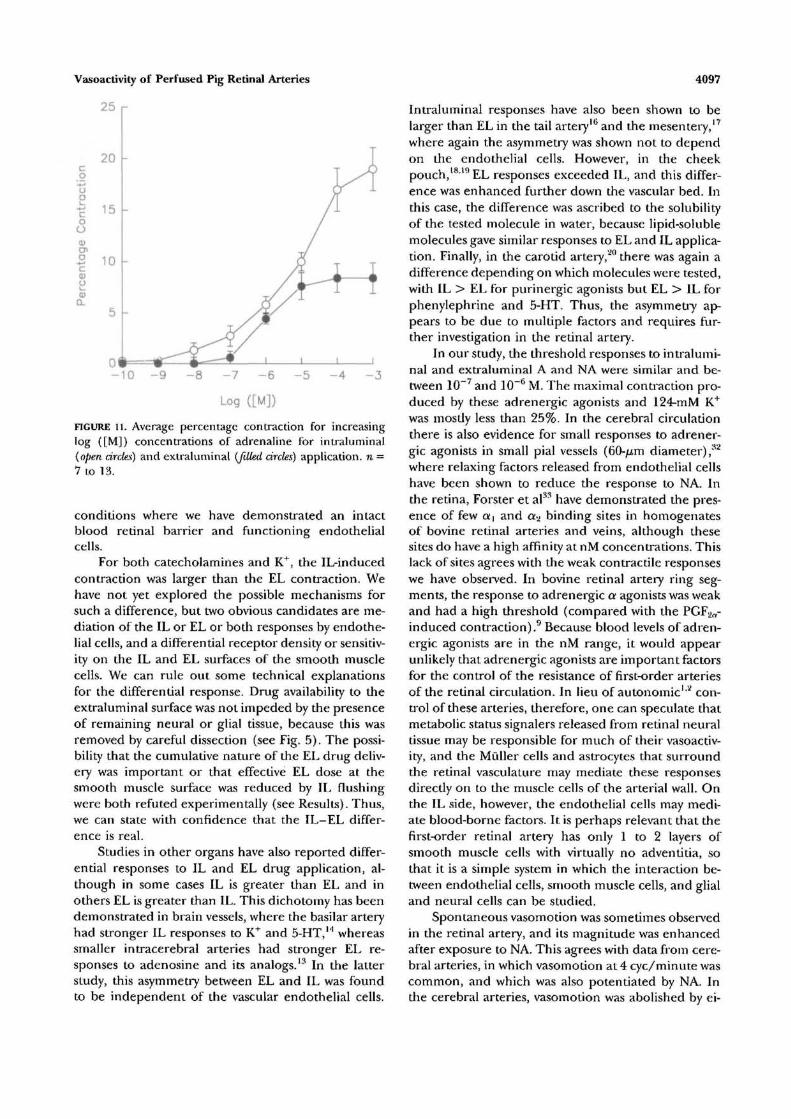
Adrenaline shows an IL-EL response pattern sim-ilar to that observed for noradrenaline. Raw data setsfor IL (top panel) and EL (bottom panel) adrenalinedelivery for a 92-//m vessel are shown in Figure 10.The contraction at 10~3-M IL was 26% of the originaldiameter, whereas the contraction with 10~3-M EL de-livery was only 8%. The averaged dose response curves(n = 7 to 13) shown in Figure 11 tell the same story,with IL delivery producing significantly larger re-sponses than EL delivery (P < 0.05) at concentrationsof 10~5 to 10~3 M. For adrenaline, the IL responses
t
8
fi
l/m
in)
it
100
502S0
200
150
100
50
012
a
4
0
-3 ioo
50250
200
^ 150
•* 100
LL
50012
I 83T 4a.
0
FIGURE 6. Graph of perfusion flow (bottom panel), P2 mm Hg {upper trace), and P4 mm Hg{lower trace) in the middle panel, and outside arterial diameter (OD /j.m, top panel) as afunction of time in minutes.
Vasoactivity of Perfused Pig Retinal Arteries 4095
110
^ too
§ 90
BO
Control
(A)
- 8 - 7 - 6 - 5
"V "\5 min
- 4
VV
rV
Control
aO
110
100
90
B0
-10
r »
(B)
- 9
I- 6
I5
5 min
- 4
I-3
1
FIGURE 8. Intraluminal dose responses to increasing doses of noradrenaline from 10 H M to10~* M (upper panel) compared with extraluminal cumulative dose responses from 10~'" Mto 10 ^ M (loxverpanel).
have a lower threshold (10 8 M) than do the EL re-sponses (10~7 M). The maximum percentage constric-tions, 19.0% ± 2.1% 1L and 8.4% ± 1.5% EL withadrenaline, are not significantly different (P > 0.05)from those for noradrenaline.
K+-induced contractions also caused differentialresponses for IL and EL application. Intraluminal in-jection of 124-mM K+ caused an average contractionof 19.0% ± 1.6% (n= 21) of the vessel's resting diame-ter, whereas EL application of the same concentration
25 r
co"uo
oo
o-+-J
cUl_0)a. 5 -
-10 - 9 - 8 - 7 - 6
Log ([M])
- 5 -4 - 3
FIGURE 9. Average percentage contraction for increasing log([M]) concentrations of noradrenaline for intraluminal(open circles) and extraluminal (filled circles) application, n =7 to 13.
caused only 5.0% ± 1.0% (n = 13). These contractionpercentages are significandy different (P < 0.001).
Blood Retinal Barrier
Using IL sodium fluorescein with fluorescent micros-copy, visualization of the perfusion of a fluorescentmarker through the vessel lumen was observed with-out leakage from either cannulation point. This indi-cated a tight cannulation widi all perfusate and drugspassing through the vessel lumen. Furthermore, move-ment of the marker across the vessel wall or betweenendothelial cells to smooth muscle cells was never ob-served. Therefore, an intact blood retinal barrier tosodium fluorescein can be assumed to exist in tfiisretinal artery preparation.
DISCUSSION
This technique of continuous IL perfusion confersseveral important advantages over the more com-monly used ring segment preparation that we3'27"30
and others5"x>i have previously used to investigate thevasoactivity of ocular arteries. A perfused artery corre-sponds more closely to the in vivo situation, allowsdifferentiation of IL and EL responses, may cause lessendothelial cell damage, especially for small vessel ofoutside diameter less than 100 //m, and is more readilyapplied to smaller vessels such as the human or por-cine retinal arteries and arterioles. Moreover, the useof a controlled perfusion technique means that theflow rate is known, and the intraluminal pressure andvessel diameter can be measured. Drug-inducedchanges in this pressure provide information aboutthe combined vascular resistance of the main vessel
4096 Investigative Ophthalmology 8c Visual Science, November 1994, Vol. 35, No. 12
and its single side branch, through which the perfu-sate exits.
Interestingly, our comparison of pressurized ves-sels with no flow and pressurized vessels with flow un-covered a strong myogenic contraction with increas-ing pressure for vessels in which there was no flow.However, with flow, but at the same pressure, the ves-sel dilates. Bevan et al22 and Kuo et al31 also observedflow-induced dilatation in isolated vessels. This effectis currently the subject of further investigations inwhich the effects of flow and intraluminal pressurecan be studied independently. It seems likely that ves-sel diameter reflects the balance of opposing forcesgenerated by mechanisms that are not well under-stood. Direct extrapolation of these in vitro findingsto the in vivo situation should be resisted because thebalance of competing vasoactive mechanisms and theadditional systemic influences are too complicated topredict.
Under our normal experimental conditionswhere flow was present, the arterial diameter was rela-tively unaffected by incremental increases in flow (Fig.6). The size of the side branch diameter determinedthe resultant intraluminal pressure. A small-diameterside branch caused high IL pressure at constant flow,whereas a large-diameter side branch resulted in lowIL pressure.
We conclude that the loss of agonist response athigh pressures and flows (Fig. 7) was a pressure effectrather than a flow effect. This conclusion is based onobservations from vessels of similar outside diameterbut with different diameter side branches. All vesselswere subjected to the same constant flow, but thosewith small diameter side branches naturally hadhigher intraluminal pressure in the main vessel than
those with larger side branches. The vessels withhigher pressures did not respond to noradrenaline,whereas those with larger side branches and lowerintravascular pressures did respond.
In parallel with these quantitative measurements,we were able to visualize clearly and observe thesmooth muscle and endothelial cells lining the lumenthrough the microscope and video camera, becauseof the thin nature of the vessel walls and the minimaladventitia (Figs. 4, 5). With change of focus, individualsmooth muscle cells and endothelial cells could beobserved. During contraction, the wall thickness wasseen to increase, leading to a proportionally largerdecrease in luminal diameter than that monitored byoutside diameter measurements.
With experience, it was possible to judge from thevisual appearance of the vessel the viability of vaso-active responses. Occasional problems with air emboliin a few experiments not included in these data re-vealed dramatic changes in cellular appearance, a pe-riod of vasospasm, and visible detachment of endothe-lial cells.
With this continuous visualization, it became ap-parent when there was some damage to the prepara-tion as the endothelial cells changed appearance, andin extreme cases detached from the smooth musclecells. These changes in appearance were confirmedby loss of vasoactivity. Thus, in conclusion, we believethat, by using this side branch preparation and associ-ated monitoring techniques, we will be able to sepa-rate vasoactive responses from side branches and fromthe main vessels in future studies.
Using this technique, we provide the first evidenceof differential responses of the retinal artery to IL andEL delivery of K+, noradrenaline, and adrenaline in
100
90
60
70
60
Control - 8 - 6 - 5 - 3 Control
jmtmmjm
100 r
90
1 80
-10 -9 - 8 - 7 - 6 - 5
(B) 5 mln
FIGURE 10. Intraluminal dose responses to increasing doses of adrenaline from 10 K M to10 ^ M (upperpanel) compared with extraluminal cumulative dose response curve from 1 0 " 'M to 10 * M (buter panel).
Vasoactivity of Perfused Pig Retinal Arteries
25 r
co
-uop"coo0)eno
•*->c<DO
- 1 0 - 9
Log ([M])
FIGURE 11. Average percentage contraction for increasinglog {[M]) concentrations of adrenaline for intraluminal(open circles) and extraluminal [filled circles) application, n =7 to 13.
conditions where we have demonstrated an intactblood retinal barrier and functioning endothelialcells.
For both catecholarnines and K+, the IL-inducedcontraction was larger than the EL contraction. Wehave not yet explored the possible mechanisms forsuch a difference, but two obvious candidates are me-diation of the IL or EL or both responses by endothe-lial cells, and a differential receptor density or sensitiv-ity on the IL and EL surfaces of the smooth musclecells. We can rule out some technical explanationsfor the differential response. Drug availability to theextraluminal surface was not impeded by the presenceof remaining neural or glial tissue, because this wasremoved by careful dissection (see Fig. 5). The possi-bility that the cumulative nature of the EL drug deliv-ery was important or that effective EL dose at thesmooth muscle surface was reduced by IL flushingwere both refuted experimentally (see Results). Thus,we can state with confidence that the IL-EL differ-ence is real.
Studies in other organs have also reported differ-ential responses to IL and EL drug application, al-though in some cases IL is greater than EL and inothers EL is greater than IL. This dichotomy has beendemonstrated in brain vessels, where the basilar arteryhad stronger IL responses to K+ and 5-HT,M whereassmaller intracerebral arteries had stronger EL re-sponses to adenosine and its analogs.13 In the latterstudy, this asymmetry between EL and IL was foundto be independent of the vascular endothelial cells.
4097
Intraluminal responses have also been shown to belarger than EL in the tail artery16 and the mesentery,17
where again the asymmetry was shown not to dependon the endothelial cells. However, in the cheekpouch,1819 EL responses exceeded IL, and this differ-ence was enhanced further down the vascular bed. Inthis case, the difference was ascribed to the solubilityof the tested molecule in water, because lipid-solublemolecules gave similar responses to EL and IL applica-tion. Finally, in the carotid artery,20 there was again adifference depending on which molecules were tested,with IL > EL for purinergic agonists but EL > IL forphenylephrine and 5-HT. Thus, the asymmetry ap-pears to be due to multiple factors and requires fur-ther investigation in the retinal artery.
In our study, the threshold responses to intralumi-nal and extraluminal A and NA were similar and be-tween 10~7 and 10~6 M. The maximal contraction pro-duced by these adrenergic agonists and 124-mM K+
was mostly less than 25%. In the cerebral circulationthere is also evidence for small responses to adrener-gic agonists in small pial vessels (60-//m diameter),32
where relaxing factors released from endothelial cellshave been shown to reduce the response to NA. Inthe retina, Forster et al33 have demonstrated the pres-ence of few a] and aa binding sites in homogenatesof bovine retinal arteries and veins, although thesesites do have a high affinity at nM concentrations. Thislack of sites agrees with the weak contractile responseswe have observed. In bovine retinal artery ring seg-ments, the response to adrenergic a agonists was weakand had a high threshold (compared with the PGFaCT-induced contraction).9 Because blood levels of adren-ergic agonists are in the nM range, it would appearunlikely that adrenergic agonists are important factorsfor the control of the resistance of first-order arteriesof the retinal circulation. In lieu of autonomic1'2 con-trol of these arteries, therefore, one can speculate thatmetabolic status signalers released from retinal neuraltissue may be responsible for much of their vasoactiv-ity, and the Muller cells and astrocytes that surroundthe retinal vasculature may mediate these responsesdirectly on to the muscle cells of the arterial wall. Onthe IL side, however, the endothelial cells may medi-ate blood-borne factors. It is perhaps relevant that thefirst-order retinal artery has only 1 to 2 layers ofsmooth muscle cells with virtually no adventitia, sothat it is a simple system in which the interaction be-tween endothelial cells, smooth muscle cells, and glialand neural cells can be studied.
Spontaneous vasomotion was sometimes observedin the retinal artery, and its magnitude was enhancedafter exposure to NA. This agrees with data from cere-bral arteries, in which vasomotion at 4 eye/minute wascommon, and which was also potentiated by NA. Inthe cerebral arteries, vasomotion was abolished by ei-

4098 Investigative Ophthalmology & Visual Science, November 1994, Vol. 35, No. 12
ther low or high pressures, leading the authors toconclude that vasomotion is a normal property of cere-bral arteries/4
In conclusion, we have developed a sophisticatedand technically demanding technique that has dem-onstrated that heterogeneity is an important factorin the retinal circulation between intraluminal andextraluminal responses. It remains to be seen whetherthis polarity of response is general for all drugs orwhether some are more active when applied extralu-minally. One might predict that vasoactive agents re-leased as metabolic signalers between tissue and theretinal vessels will be most effective on the EL side,but that remains to be determined. The data pre-sented here demonstrate that this technique is capableof providing reliable and reproducible data and thatit has considerable potential to enhance our under-standing of retinal vascular control. Indeed, it is al-ready clear that the first-order retinal circulation dif-fers substantially from the ophthalmociliary artery inits pharmacologic responses, and it is likely that thisheterogeneity in response continues down the vascu-lar pathway. Thus, for a complete understanding ofthe sites and actions of drugs in the retinal circulationin health and disease, it is essential that all regions ofthe vasculature from conduit artery to capillaries andveins be studied. Moreover, the perfused retinal arteryoffers an exciting system to study more fundamentalquestions interrelating smooth muscle and endothe-lial cell function for vascular physiologists, because itis a vessel with a simple wall structure, only 1 to 2layers of smooth muscle cells, an endothelial cell layer,and no confounding autonomic nerve endings.
Key Words
vasoactivity, perfused artery, pig, retina, agonists
Acknowledgments
The authors thank Dean Darcey, Michael Brown, and PeterBurrows for their excellent technical assistance.
References
1. Laties AM. Central retinal artery innervation: Absenceof adrenergic innervation to the intraocular branches.Arch Ophthalmol 1967; 77:405-409.
2. Ye X, Laties AM, Stone RA. Peptidergic innervationof the retinal vasculature and optic nerve head. InvestOphthlamol Vis Sri. 1990;31:l731-l737.
3. Yu D-Y, Su E-N, Alder VA, Cringle SJ, Mele EM.Pharmacological and mechanical heterogeneity of catisolated ophthalmociliary artery. Exp Eye Res.1992; 54:347-359.
4. Thuranszky K. Der Blutkreislauf der Netshaut. Budapest:Ungarischen Akademie der Wissenshaft; 1957.
5. Hoste AM, Andries LJ. Contractile responses of iso-lated bovine retinal microarteries to acetycholine. In-vest Ophthalmol Vis Sri. 1991;32:1996-2005.
6. Benedito S, Prieto D, Nielsen PJ, Nyborg NCB. Role ofthe endothelium in acetylcholine-induced relaxationand spontaneous tone of bovine isolated retinal smallarteries. Exp Eye Res. 1991;52:575-579.
7. Nielsen PJ, Nyborg NCB. Contractile and relaxing ef-fects of arachidonic acid derivatives on isolated bovineretinal resistance arteries. Exp Eye Res. 1990; 50:305-311.
8. Nielsen PJ, Nyborg NCB. Calcium antagonist-inducedrelaxation of the prostaglandin-F2a response of iso-lated calf retinal resistance arteries. Exp Eye Res.1989;48:329-335.
9. Nielsen PJ, Nyborg NCB. Adrenergic responses in iso-lated bovine retinal resistance arteries. Int. Ophthalmol.1989;13:103-107.
10. Hoste AM, Boels PJ, Andries LJ, Brutsaert DL, De LaeyJ. Effects of beta-antagonists on contraction of bovineretinal microarteries in vitro. Invest Ophthalmol Vis Sri.1990;31:1231-1237.
11. Nyborg NCB, Prieto D, Benedito S, Nielsen PJ. Endo-thelin-1-induced contraction of bovine retinal smallarteries is reversible and abolished by nitrendipirie.Invest Ophthalmol Vis Sri.. 1991;32:27-31.
12. Nyborg NCB, Nielsen PJ. Thrombin contracts isolatedbovine retinal small arteries in vitro. Invest OphthalmolVis Sri. 1990;31:2307-2313.
13. Ngai AC, Winn HR. Effects of adenosine and its ana-logues on isolated intracerebral arterioles. Circ Res.1993; 73:448-457.
14. Ogawa R, Ohta T, Tsuji M, Mori M. Role of the endo-thelium on extraluminal and intraluminal vasoactivemechanisms in the perfused rabbit basilar artery. Neu-rolRes. 1993; 15:154-159.
15. HeadrickJP, Northington FJ, Hynes MR, MatherneGP, Berne RM. Relative responses to luminal and ad-ventitial adenosine in perfused arteries. AmJ Physiol.1992;263:H1437-H1446.
16. Mazmanian GM, Baudet B, Pannier-Poulain C, et al.Pressor responses of rat isolated tail arteries to con-tractile stimulation after methylene blue treatment:Effect of adventitial versus intimal entry. / Vase Res.1993; 30:250-256.
17. Tesfamariam B, Halpern W. Asymmetry of responsesto norepinephrine in perfused resistance arteries. EurJ Pharmacol. 1988; 152:167-170.
18. Lew MJ, Duling BR. Access of blood-borne vasocon-strictors to the arteriolar smooth muscle. / Vase Res.1992;29:341-346.
19. Matsuki T, Hynes MR, Duling BR. Comparison of con-duit vessel and resistance vessel reactivity: Influenceof intimal permeability. AmJPhysiol. 1993;264:H1251-H1258.
20. Kaul S, Waak BJ, Heistad DD. Asymmetry of vascularresponses of perfused rabbit carotid artery to intralu-minal and abluminal vasoactive stimuli. / Physiol.1992;458:223-234.
21. Jackson WF, Duling BR. The oxygen sensitivity of ham-ster cheek pouch arterioles. Cire Res. 1983;53:515-525.
22. Bevan JA, Joyce EH, Wellman GC. Flow-dependent

Vasoactivity of Perfused Pig Retinal Arteries 4099
dilation in a resistance artery still occurs after endo-thelial removal, drc Res. 1988;63:980-985.
23. Chonko AM, Irish JM III, Welling DJ. Microperfusionof isolated tubules. In: Martinez-Maldonado M, ed.Methods in Pharmacology: Vol. 4B, Renal Pharmacology.
New York: Plenum Press; 1978:221-258.24. Duling BR, Rivers RJ. Isolation, cannulation and per-
fusion of microvessels. In: Baker CH, Nastuk WL, eds.Microcirculatory Technology. New York: Academic Press;1986.
25. Murta JN, Cunha-Vaz JG, Sabo CA, Jones CW, LaskiME. Microperfusion studies on the permeability ofretinal vessels. Invest Ophthalmol Vis Sci. 1990; 31:471-480.
26. Riva CE, Grunwald JE, Sinclair SH, Petrig BL. Bloodvelocity and volumetric flow rate in human retinalvessels. Invest Ophthalmol Vis Sci. 1985;26:1124-1132.
27. Yu D-Y, Alder VA, Cringle SJ. In vitro characterizationof the mechanical properties of canine ophthalmicartery. Exp Eye Res. 1990;51:729-734.
28. Yu D-Y, Alder VA, Su E-N, Mele EM, Cringle SJ,
Morgan WH. Agonist response of human isolated pos-terior ciliary artery. Invest Ophthalmol Vis Sci.1992;53:48-54.
29. Alder VA, Su E-N, Yu D-Y, Cringle SJ. Oxygen reac-tivity of the feline isolated ophthalmociliary artery.Invest Ophthalmol Vis Sci. 1993; 34:49-57.
30. Su E-N, Yu D-Y, Alder VA, Cringle SJ. Effects ofextracellular pH on agonist-induced vascular tone ofthe cat ophthalmociliary artery. Invest Ophthalmol VisSci. 1994;35:998-1007.
31. Bauknight GC, Faraci FM, Heisted DD. Endothelium-derived relaxing factor modulates noradrenergic con-striction of cerebral arterioles in rabbits. Stroke.1992;23:1522-1526.
32. Forster BA, Ferrari-Dileo G, Anderson DR. Adrenergicalphal and alpha2 binding sites are present in bovineretina and retinal blood vessels. Invest Ophthalmol VisSci. 1987; 28:1741-1746.
33. Fujii K, Heisted DD, Faraci FM. Vasomotion of basilararteries in vivo. AmJPhysiol. 1990;258:H1829-H1834.
34. Kuo L, Davis MJ, Chilian WM. Am J Physiol.1990;259:H1036-H1070.




![Investigation of factors affecting fluorometric quantitation of cytosolic [Ca2+] in perfused hearts](https://img.pdfslide.net/doc/110x75/634ddcabd38be601b805ed3b/investigation-of-factors-affecting-fluorometric-quantitation-of-cytosolic-ca2.jpg)






















