Embed Size (px)
Citation preview

Immunology, 1973, 25, 323.
Guinea-pig Reaginic Antibody
I. ISOLATION AND PURIFICATION OF AN ANTIBODYPROTEIN WITH CHARACTERISTICS OF IgE
R. A. MARGNI AND SILVIA E. HAJOS
Department of Microbiology, Chair of Immunology,Faculty of Pharmacy and Biochemistry, Buenos Aires University,
Junin 956, Buenos Aires, Argentina
(Received 8th January 1973; acceptedfor publication 7th February 1973)
Summary. The methods for isolation and purification of a guinea-pig serumprotein with homocytotropic antibody activity and characteristics of IgE aredescribed.By precipitation in the equivalence zone or immunoadsorption and chromato-
graphy on DEAE-cellulose, we isolated an homocytotropic antibody, that was notable to give a precipitin line when it was reacted directly with the antigen. It wascapable of sensitizing guinea-pig skin for PCA after a latent period of 24 48 hoursbut not after 3 hours; it was sensitive to treatment with mercaptoethanol. It hadantigenic determinants present in the other guinea-pig immunoglobulins andparticular antigenic determinants.
All these properties make us believe that this protein belongs to an immuno-globulin different from yi and similar to the reaginic antibody (IgE) described inother species.
INTRODUCTION
A great number ofhomocytotropic and heterocytotropic antibodies have been identifiedin many mammalian species (Benacerraf, Ovary, Block and Franklin, 1963; Fahey,Wunderlich and Mishell, 1964; Ovary, Barth and Fahey, 1965; Ishizaka, Ishizaka andHornbrook, 1966, 1970; Zvaifler and Becker, 1966; Mota and Peixoto, 1966; Mota,1967; McVeigh and Voss, 1969; Van Ess, Dan Harink and Pondman, 1970; Stechschulte,C. Orange and Austen, 1970).
In the mouse the simultaneous presence of two different homocytotropic antibodies hasbeen reported, one of them located in the IgG-y1 subclass and the other in an immuno-globulin with similar characteristics to IgE human reaginic antibody (Mota and Peixoto,1966; Mota, 1967).In the guinea-pig the homocytotropic antibody corresponds to the IgG-y1 but recently
the existence of an anaphylactic antibody similar to the one described in the mouse hasbeen demonstrated although it has not been isolated in a purified state yet (Van Ess,Dan Harink and Pondman, 1970; Mota and Perini, 1970; Parish, 1970; Levine, Changand Vaz, 1971; Dobson, Norset and Soulsby, 1971).
In the course of previous investigations with guinea-pig non-precipitating antibodies*(Margni and Hajos, 1973) a protein with IgE characteristics was found.
323

R. A. Margni and Silvia E. Hajos
The present study has been developed to attempt the isolation and purification of thisguinea-pig antibody.
MATERIALS AND METHODS
Dinitrophenyl derivativesDinitrophenyl human y-globulin (DNP-HGG) and dinitrophenyl bovine serum albumin
(DNP-BSA) were prepared as described by Russell Little and Eisen (1967). Theycontained thirty-eight and thirty-two haptenic groups per mole of protein, respectively.
AntiseraPools of sera with anti-DNP homocytotropic activity (PCA reaction after a 72 hours
sensitization latent period) obtained from 150-200 immunized random-bred guinea-pigsin each case, were analysed. The guinea-pig sera were obtained from animals inoculatedwith DNP-HGG mixed with Freund's complete adjuvant or Bordetella pertussis.Groups of animals were injected intramuscularly in both hind legs with 0-5 ml of
DNP-HGG 0.1 per cent emulsified with an equal amount of Freund's complete adjuvant.A week after the first antigen dose the animals received another injection and after 3weeks of resting they were boosted and bled 10-12 days later.
Other groups of guinea-pigs were inoculated with DNP-HGG in saline or with DNP-HGG in saline and simultaneous injection intraperitoneally of 1 ml of heat-killed Bordetellapertussis phase I (15 x 109 bacteria per ml). The schedule used was the same indicated forthe first group.
Isolation and purification of antibodyThe purification was made in two different ways. In the first case, total antibodies were
precipitated with DNP-BSA in the equivalence zone, dissociated with 0-1 M dinitrophenol(DNP-OH) and applied to a DEAE-cellulose and Amberlite IRA-400 column, accordingto Eisen, Gray, Russell Little and Sims (1967).
In the second case, the serum was passed through an immunoadsorbent column. Asimmunoadsorbent, DNP-BSA polymerized with glutaraldehyde was used (Avrameas andTernynck, 1969; Margni and Binaghi, 1972). For preparation of the immunoadsorbentBSA was first polymerized with glutaraldehyde (10 ml BSA 4 per cent in 0 2 M aceticbuffer, pH 5, plus 2 ml 2 -5 per cent glutaraldehyde in the same buffer, 24 hours in the cold).
After polymerization the BSA was broken with a glass homogenizator into smallpieces, washed, taken in 0 25 per cent Na2 CO3 (1 ml/10 mg BSA) and conjugated with10 per cent fluordinitrobenzene (FDNB) in dioxane. The quantity of FDNB employedwas 25 per cent of the weight of BSA. After 2 hours at room temperature with constantstirring and overnight in the cold, the polymerized DNP-BSA was washed and used asimmunoadsorbent.The antibodies were dissociated with 0 1 M DNP-OH and purified by passage through
an Amberlite IRA-400 column, eluting with 0-15M NaCL 0-01 M phosphate, pH 7 6.The isolation of Y2, yi and reaginic antibody was carried out by passage through a
DEAE-cellulose column equilibrated in 0-01 M phosphate buffer, pH 7 6, and elutingwith the same buffer until total isolation of Y2. Then a gradient of elution with 0 01 Mphosphate, pH 7 -6, as the starting buffer and 0 3 M phosphate, pH 7 -6, as the final one wasapplied. An LKB fraction collector was used.
324

Guinea-pig Reaginic Antibody. IThe protein peaks corresponding to Y2, yi and reaginic antibody were recycled in
DEAE-cellulose. The recycling of Y2 was performed with an elution gradient of 0 003/0015 M, pH 7 6. The recycling of yi was done similarly with 0-01 M phosphate, pH 7 6,as the starting buffer and 0 05 M phosphate, pH 7 6, as the final one. For reaginic antibody,recycling was performed beginning with 0 03 M phosphate, pH 7 -6, and ending with 0 3 Mphosphate, pH 7 6.
ImmunoelectrophoresisIt was performed by the micro-method of Scheidegger (1955) using an LKB equipment.
1 25 per cent agar in 0-025 M veronal buffer, pH 8-2, was used.
ImmunodiffusionDouble diffusion in agar was carried out by Ouchterlony's method (Ouchterlony,
1958).
Cellulose acetate electrophoresisGelatinized cellulose acetate (Cellogel, Chemetron, Milan) and the method that was
described in other papers was used (Margni et al., 1970a, b; Heer and Margni, 1971).
Acrylamide gel electrophoresisWas made according to Orstein and Davis (1962) with a minor modification. To obtain
a medium pore gel, solution C was prepared with 20 g acrylamide and 1 25 g bis-acryla-mide.
HaemagglutinationDinitrophenylated sheep red cells were used according to Bullock and Kantor (1965)
with a minor modification according to Dr Binaghi's personal suggestion. 2,4-Dinitro-fluorbenzene (FDNB) was used instead of F2 DNB. As the reaction is slow, it is necessaryto carry it out in an hypertonic medium, in order to avoid passage of a large proportion ofthe FDNB into cells, thus causing poor labelling of the membrane lysines. The problemwas solved by replacing the buffer No. 1 of Bullock and Kantor by a similar buffer inwhich the NaCl concentration was 0 3 M instead of0* 15 M. Once the red cells were labelled,they were washed with the original isotonic buffer.
Serial two-fold dilutions of 10 pg N Ab/ml solution were made in 0 5 ml of saline buffer.0-05 ml of a 2 per cent suspension of dinitrophenylated sheep red cells were added to eachtube. The patterns of haemagglutination were read after the tubes had been left 15minutes at 370 and overnight at room temperature. Tubes with non-dinitrophenylatedsheep red cells, without antibody and tubes with guinea-pig yi tetanus anti-toxin wereincluded as controls.
Passive cutaneous anaphylaxis (PCA)This was performed according to Ovary (1952). The guinea-pigs were inoculated
intradermally with 0-1 ml of the antibody solutions. One milligram of DNP-BSA/0.5 mlmixed with an equal volume of 0 5 per cent Evans' blue was used as challenge dose. Itwas inoculated at different times.
Spots below a diameter of 0 5 cm were considered dubious reactions, while the ones
325

R. A. Margni and Silvia E. Hajos
between 0 5-1 cm, I90-1-5 cm, 1 -5-2 cm and more than 2 cm were considered as positive(+), (+ +), (+ + +) and (+ + + +) respectively.
Mast cell degranulationThis was carried out according to Mathov and Grinstein (1969). Peritoneal cells of
non-immune guinea-pigs were placed on slides and incubated with the antibody (1 6 YgN Ab/ml) at 40 during 30 minutes; washed; incubated with 0 05 per cent DNP-BSA at40 during 20 minutes; washed; fixed with methanol, acetic acid and water (50:3:50) andstained with thionine 1 per cent in water.
AntiseraThe rabbit anti-guinea-pig serum was prepared by repeated inoculation of 1 ml of
guinea-pig serum emulsified with an equal volume of Freund's complete adjuvant. Theanimals received a weekly injection for 3 weeks and another one 3 weeks later. After aweek they were bled.The anti-reaginic serum was obtained by injecting rats with 250 pg of purified reaginic
antibody in 0 5 ml of 0-15 M NaCl mixed with an equal volume of Freund's completeadjuvant. Another injection was given 7 days later and after 3 weeks of resting the animalswere boosted in a similar way. Between the sixth and tenth day following the boosting,the animals were bled. One part of the serum was used whole and another part wasabsorbed with guinea-pig y1 and Y2 to make it monospecific. For the absorptions a series oftubes containing equal volumes of immune rat serum were mixed with increasing quanti-ties of a mixture of equal parts (w/v) ofguinea-pigyiY2 andY3. The tubes were incubated at370 for 60 minutes and at 40 for 24 hours. After centrifugation, the supernatants that didnot react with guinea-pig yV and V2, were passed through a column containing poly-merized new born guinea-pig sera lacking reaginic activity and checked by immunodiffu-sion and immunoelectrophoresis with Vt, Y2, purified reaginic antibody, new born guinea-pig sera lacking reaginic activity, and non-immunized guinea-pig sera.The monospecific anti-reaginic serum used was the one that only reacted with the purified
reaginic antibody.The polymerized new born guinea-pig serum used for absorption was prepared by the
method described for polymerization of DNP-BSA. Phosphate buffer 0f1 M, pH 6 2, wasemployed.
Reduction and alkylationThe purified guinea-pig antibody was treated for 1 hour at room temperature with
mercaptoethanol until a 0-1 M final concentration followed by alkylation with iodoaceta-mide until 0-2 M final concentration for 30 minutes at 4°. The material was then dialysedagainst saline phosphate.
Storage and stability ofpurified guinea-pig reaginic antibodyPools of purified reaginic antibody were kept frozen at -200. Repeated freezing and
thawing was avoided. Significant changes were observed in their capacity to react withpolymerized DNP-BSA and to mediate PCA after a 48-72-hour sensitization period.
Radio-labelling of antibodyLabelling with 125I was performed with chloramine T, as described by McConahey and
Dixon (1966).
326
Guinea-pig Reaginic Antibody. I
RESULTS
The isolation and purification of guinea-pig reaginic antibody with DNP specificitywas possible by the two methods described above.When the precipitin reaction was done in the equivalence zone, another different anti-
body with electrophoretic mobility faster than Vi coprecipitated with yi and Y2* Whensmall amounts ofantigen (approximately 1 /20ofthe antigennceded foroptimal precipitation)were added successively, this antibody was included in the precipitate at minimal concen-trations and remains in the supernatant. This antibody was also obtained by passing theantisera through an immunoabsorbent column.
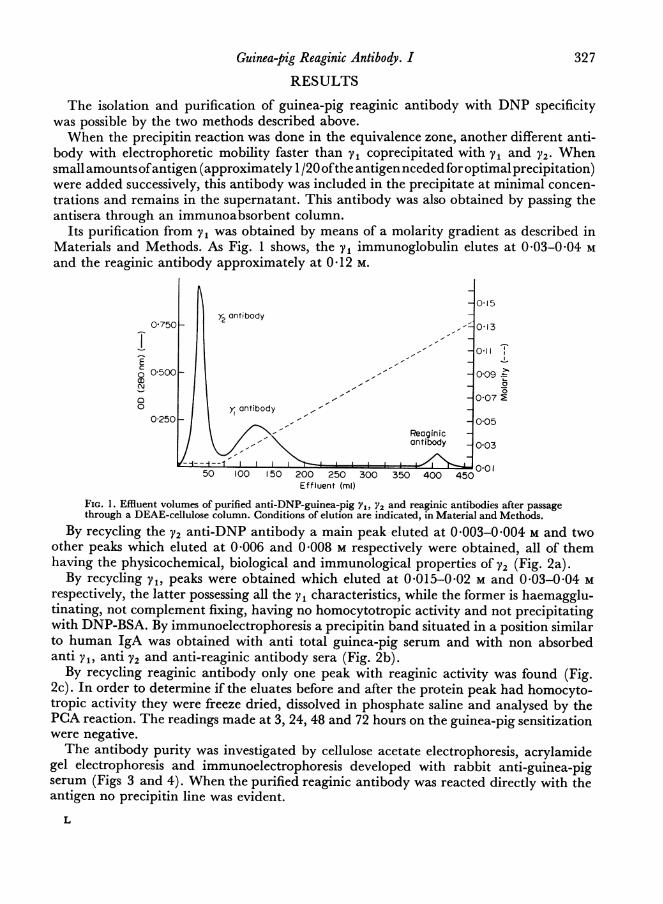
Its purification from yV was obtained by means of a molarity gradient as described inMaterials and Methods. As Fig. 1 shows, the Vi immunoglobulin elutes at 0 03-0-04 Mand the reaginic antibody approximately at 0-12 M.
0-1502 antibody
0 7502 0.-1 3
Co 0500 009
0o 0*07 E0 l antibody -
0250 0- 005Reaginicantibody -003
0-0150 100 150 200 250 300 350 400 450
Effluent (ml)
FIG. 1. Effluent volumes of purified anti-DNP-guinea-pig Y1, Y2 and reaginic antibodies after passagethrough a DEAE-cellulose column. Conditions of elution are indicated, in Material and Methods.
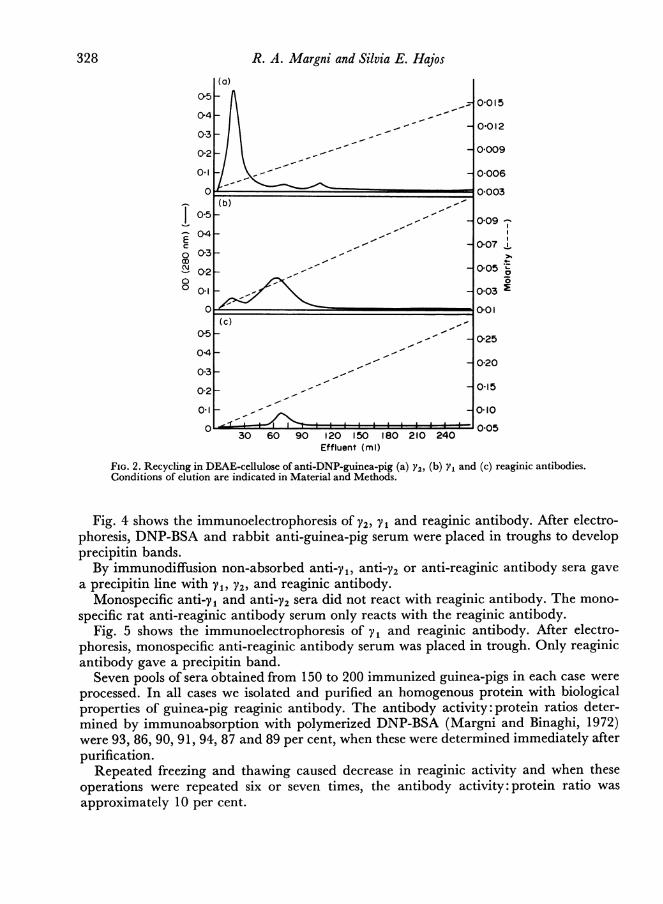
By recycling the V2 anti-DNP antibody a main peak eluted at 0 003-0 004 M and twoother peaks which eluted at 0 006 and 0 008 M respectively were obtained, all of themhaving the physicochemical, biological and immunological properties of V2 (Fig. 2a).By recycling yV, peaks were obtained which eluted at 0-015-0-02 M and 0-03-0-04 M
respectively, the latter possessing all the yV characteristics, while the former is haemagglu-tinating, not complement fixing, having no homocytotropic activity and not precipitatingwith DNP-BSA. By immunoelectrophoresis a precipitin band situated in a position similarto human IgA was obtained with anti total guinea-pig serum and with non absorbedanti y1, anti V2 and anti-reaginic antibody sera (Fig. 2b).By recycling reaginic antibody only one peak with reaginic activity was found (Fig.
2c). In order to determine if the eluates before and after the protein peak had homocyto-tropic activity they were freeze dried, dissolved in phosphate saline and analysed by thePCA reaction. The readings made at 3, 24, 48 and 72 hours on the guinea-pig sensitizationwere negative.The antibody purity was investigated by cellulose acetate electrophoresis, acrylamide
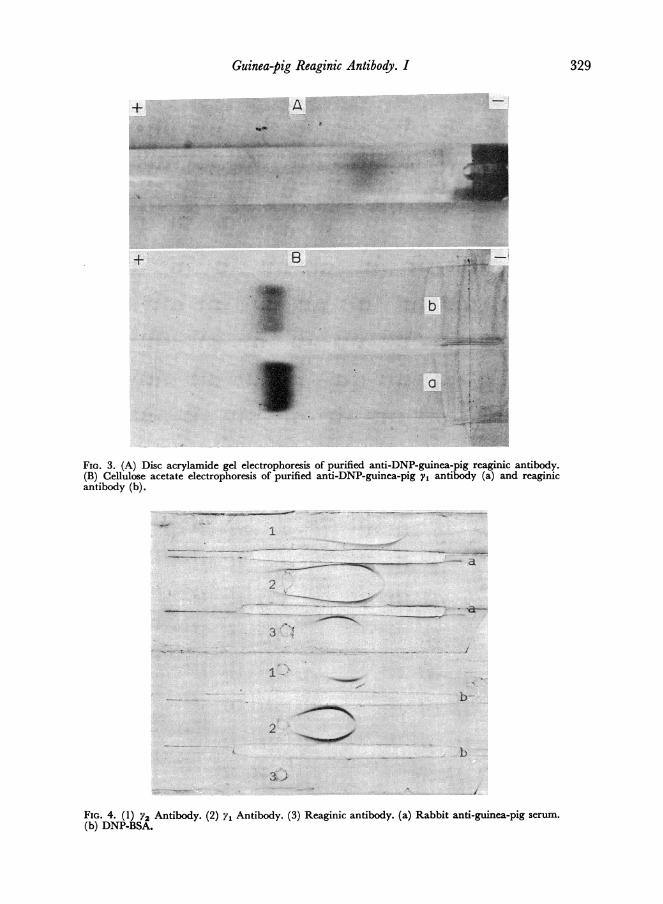
gel electrophoresis and immunoelectrophoresis developed with rabbit anti-guinea-pigserum (Figs 3 and 4). When the purified reaginic antibody was reacted directly with theantigen no precipitin line was evident.
L
327
R. A. Margni and Silvia E. Hajos
0-5 -0097
C 0-07o 03-3
0 _ , - '4-OSc0-2 -5*
00.1 0-03
C _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ 01-01
(c)0 5_ ,- _ 0-25
0-4 _,4- 0-20
0-3 _
0-2 _ __ - ~ _0-15
0-2 --0-1
C 0-0530 60 90 120 150 180 210 240
Effluent (ml)
FIG. 2. Recycling in DEAE-cellulose of anti-DNP-guinea-pig (a) Y2, (b) Y1 and (c) reaginic antibodies.Conditions of elution are indicated in Material and Methods.
Fig. 4 shows the immunoelectrophoresis of V2, Vi and reaginic antibody. After electro-phoresis, DNP-BSA and rabbit anti-guinea-pig serum were placed in troughs to developprecipitin bands.By immunodiffusion non-absorbed anti-y1, anti-Y2 or anti-reaginic antibody sera gave
a precipitin line with Vy, V2, and reaginic antibody.Monospecific anti-V1 and anti-Y2 sera did not react with reaginic antibody. The mono-
specific rat anti-reaginic antibody serum only reacts with the reaginic antibody.Fig. 5 shows the immunoelectrophoresis of yV and reaginic antibody. After electro-
phoresis, monospecific anti-reaginic antibody serum was placed in trough. Only reaginicantibody gave a precipitin band.
Seven pools of sera obtained from 150 to 200 immunized guinea-pigs in each case wereprocessed. In all cases we isolated and purified an homogenous protein with biologicalproperties of guinea-pig reaginic antibody. The antibody activity: protein ratios deter-mined by immunoabsorption with polymerized DNP-BSA (Margni and Binaghi, 1972)were 93, 86, 90, 91, 94, 87 and 89 per cent, when these were determined immediately afterpurification.
Repeated freezing and thawing caused decrease in reaginic activity and when theseoperations were repeated six or seven times, the antibody activity: protein ratio wasapproximately 10 per cent.
328
Guinea-pig Reaginic Antibody. I
FIG. 3. (A) Disc acrylamide gel electrophoresis of purified anti-DNP-guinea-pig reaginic antibody.(B) Cellulose acetate electrophoresis of purified anti-DNP-guinea-pig y, antibody (a) and reaginicantibody (b).
FIG. 4. (1) Y2 Antibody. (2) y1 Antibody. (3) Reaginic antibody. (a) Rabbit anti-guinea-pig serum.
(b) DNP-BSA.
329

R. A. Margni and Silvia E. Hajos
FIG. 5. Immunoelectrophoresis. (4) Reaginic antibody. (5) Yi Antibody. (c) Monospecific rat anti-reaginic serum.
yV Antibody gives a PCA reaction after a 3-hour sensitization period. For reaginicantibody the PCA reaction is negative after a 3-hour latent period, becomes positive at24-48 hours and persists at passively sensitized skin sites for some days. The PCA reactionis negative if the reaginic antibody is previously reduced with mercaptoethanol.When 2 ml of 8 pg N Ab/ml of protein with reaginic activity and antibody activity:
protein ratio not less than 90 per cent were mixed with polymerized monospecific ratanti-reaginic antibody serum (5 ml ofserum plus 3 ml phosphate 0 1 M, pH 6 *2, polymerizedwith glutaraldehyde), the supernatant did not give PCA after a 72-hour sensitizationperiod and the passive haemagglutination and mast-cell degranulation were negative.When 2 ml (1 -6 pg N Ab/ml) of the same protein labelled with 125I was mixed withpolymerized monospecific rat anti-reaginic antibody serum, the immunoabsorbent boundthe radioactivity.No absorption was obtained with polymerized normal rat serum.
DISCUSSION
There is no doubt that the guinea-pig produces two different homocytotropic antibodies,perfectly separable by DEAE-cellulose. The y 1 antibody elutes with 0 -03-0 -04 M phosphatebuffer, pH 7 *6, while the reaginic antibody elutes at the same pH but at 0 * 12 M concentra-tion.As it co-precipitates with yV and V2 it is possible to isolate it from the total antibodies
precipitated in the equivalence zone. If the antigen is added in small amounts (1/20 of theantigen necessary for optimal precipitation) the reaginic antibody does not precipitate, itremains in the supernatant and can be isolated by passage through an immunoadsorbent.
According to the electrophoretic and immunoelectrophoretic analyses it appears thatthere are some differences in their electrophoretic mobility, one with IgG-y1 mobilityand the reaginic antibody with a slightly faster one.The anti-y1, anti-y2 and anti-reaginic sera without any absorption give a precipitin line
with yV, V2 or reaginic antibody. After being absorbed with yV, V2 and polymerized newborn guinea-pig serum, rat anti-reaginic serum only reacts with the reaginic antibody.That shows antigenic relations between these proteins and the presence in the reaginicantibody of particular antigenic determinants. It has the antigenic characteristics of animmunoglobulin.
Although it is an homocytotropic antibody it differs from y in its skin sensitizing latentperiod. The PCA reaction is negative after a 3-hour latent period and is positive after
330

Guinea-pig Reaginic Antibody. I 331
24-48 hours latent period. On the other hand, yi is capable of inducing PCA after a 3-hour latent period but becomes negative after 48 hours.The reaginic antibody is a labile protein; it loses activity with repeated freezing and
thawing. The skin sensitizing activity is completely destroyed after reduction withmercaptoethanol.
It may be agreed that the isolated globulin is IgA contaminated with reaginic antibody,as in man. If that were so, the precipitin line would be due to IgA, and the reaginic activityto the IgE protein would not be detected in the gel diffusion system. This is probably notthe case since the peak corresponding to the reaginic antibody continues to be homo-geneous after recycling. Eluates obtained after recycling corresponding to the anteriorand posterior parts of the protein peak, without absorbance at 280 nm showed no homo-cytotropic activity. Besides, the recycling of the protein fraction corresponding to Vy, givesat 0 015-0-02 M phosphate a new protein peak with antibody activity having some of thecharacteristics of IgA, which does not react with anti-reaginic monospecific serum. Thisantibody is at present under study.On the other hand the PCA supernatant did not give a reaction after a 72-hour sensi-
tization period and the passive haemagglutination and mast-cell degranulation werenegative when this protein was mixed with polymerized monospecific anti-reaginicantibody serum. If the purified protein with reaginic activity is labelled with 125I, all theradioactivity is retained by the immunoadsorbent. It shows that there is a quantitativerelation between homocytotropic activity and protein content.We consider that the purified isolated protein, present in the guinea-pig serum at
minimal concentrations, is the reaginic antibody and that it belongs to a different immuno-globulin class, different from yV and similar to human IgE and other reaginic antibodiesfrom other mammalian species.The possibility that the isolated protein is a non-precipitating yI antibody was excluded.
In our previous studies of guinea-pig non-precipitating antibodies (Margni and Hajos,1973) we found that the guinea-pig yV non-precipitating antibody had the same physico-chemical and biological properties as the yV precipitating one.The failure to isolate it until now might be due to the use ofthe method usually employed
for isolation of guinea-pig antibodies. By DEAE-cellulose step-wise elution with M 0-01,pH 7 -6 and M 0 1, pH 7 -6 phosphate, it is only possible to separate V2 and yI immunoglo-bulins. In these conditions, the reaginic antibody is not eluted; it remains bound at theDEAE-cellulose. If M 0 2, pH 7-6, phosphate is added, elution is possible.On the other hand, the presence of reaginic antibody in the immunized guinea-pig
serum is directly related to the schedule of immunization used and the time of bleedingthe animals.
ACKNOWLEDGMENTSSupported with part of the Grants: Carp 1/71, Facultad de Farmacia y Bioquimica de
la Universidad de Buenos Aires; 3886/69 Consejo Nacional de Investigaciones Cientificasy Tecnicas de la Republica Argentina; CQ-69 Cerveceria y Malteria Argentina Quilmes.
REFERENCESAVRAMEAS, S. and TERNYNCK, T. (1969). 'The Cross-
linking of proteins with glutaraldehyde and its usefor the preparation of immunoadsorbents.' Immuno-chemistry, 6, 53.
BENACERRAF, B., OvARY, Z., BLOCH, K. and FRANKLIN,E. (1963). 'Properties of guinea pig 7S antibodies.I. Electrophoretic separation of two types of guineapig 7S antibodies.' J. exp. Med., 117, 937.

332 R. A. Margni and Silvia E. HajosBINAGHI, R. (1966). 'Production of 7S immunoglobu-
lins in immunized guinea pigs.'_3. Immunol., 97, 159.BULLOCK, W. E. and KANTOR, F. S. (1965). 'Hemag-
glutination reactions of human erytrocytes con-jugated covalently with dinitrophenyl groups.' J.Immunol., 94, 317.
DOBSON, C., NORSETH, D. J. and SOULSBY, E. J. L.(1971). 'Immunoglobulin E-type antibodies inducedby Ascaris Suun infections in guinea pigs.' 3.Immunol., 106, 128.
EISEN, H. N., GRAY, W., RUSSELL LITTLE, J. andSIMs, E. S. (1967). Methods in Immunology andImmunochemistry. (Ed. by C. A. Williams andM. W. Chase). p. 353. Academic Press, Inc. NewYork.
FAHEY,J. L., WUNDERLICH,J. and MISHELL, R. (1964).'The immunoglobulins of mice. II. Two subclasses ofmouse 7S Y2 globulins; Y2a and Y2b globulins.' J.exp. Med., 120, 243.
HEER, E. E. and MARGNI, R. A. (1971). Electrophoresise Inmunoelectroforesis (Ed. by G. Fernandez), BuenosAires.
ISHIZAKA, K., ISHIZAKA, T. and HORNBROOK, M. (1966).'Physicochemical properties of human reaginicantibody. IV. Presence of a unique immunoglobulinas a carrier ofreaginic antibody.'7. Immunol., 97, 75.
ISHIZAKA, K., ISHIZAKA, T. and HORNBROOK, M.(1970). 'A unique rabbit immunoglobulin havinghomocytotropic antibody activity.' Immunochemistry,7, 515.
LEVINE, B. B., CHANG, H. and VAz, N. M. (1971).'The production of hapten-specific reaginic anti-bodies in the guinea pig.' 3. Immunol., 106, 29.
MARGNI, R. A., ACERBO, R. O., HEER, E. E. andHAjos, S. E. (1970a). 'The importance of immuno-chemical analyses in the differential diagnosis ofmicroglobulinic myelomas.' Clin. Chim. Acta, 28, 451.
MARGNI, R. A., HEER, E. E., ACERBO, E. O., HAjos,S. E., BELIVEAU, M. and BOBBI, M. E. C. DE. (1970b).'Immunochemical and genetic studies in two bis-albuminemic families.' Clin. Chim. Acta, 29, 219.
MARGNI, R. A. and BINAGHI, R. (1972). 'Purificationand properties ofnon precipitating rabbit antibodies.'Immunology, 22, 557.
MARGNI, R. A. and HAjos, S. E. (1973). 'Physico-chemical and biological properties of purified antiDNP guinea pig non-precipitating antibodies.' Im-munology, 24, 435.
MATHOV, E. and GRINSTEIN, M. (1969). 'Degranula-cion de los mastocitos en el peritoneo de cobayodurante el choque anafilactico.' Alergia, 26, 179.
MCCONAHEY, P. J. and DIXON, F. J. (1966). 'A methodof trace iodination of proteins for immunologicstudies'. Inc. Arch. Allergy, 29, 185.
MCVEIGH, T. and Voss, E. (1969). 'Properties ofpurified mouse homocytotropic antibody.' J. Im-munol., 103, 1349.
MOTA, I. and PEIXoTo,J. M. (1966). 'A skin-sensitizingand thermolabile antibody in the mouse.' LifeSciences, 5, 1723.
MOTA, I. (1967). 'Biological characterization of mouseearly antibodies.' Immunologv, 12, 343.
MOTA, I. and PERINI, A. (1970). 'A heat labilemercaptoethanol susceptible homocytotropic anti-body in the guinea pig.' Life Science, 9, 923.
ORSTEIN, L. and DAVIS, J. (1962). Disc Electrophoresis.Distillation products industries. Rochester, N.Y.
OUCHTERLONY, 0. (1958). 'Diffusion-in-gel-methodsfor immunological analyses.' Progr. Allergy, 5, 1.
OVARY, Z., BARTH, W. F. and FAHEY, J. (1965). 'Theimmunoglobulins of mice. III. Skin sensitizingactivity of mouse immunoglobulins.'J. Immunol., 94,410.
OVARY, Z. (1952). 'Cutaneous anaphylaxis in thealbino rat.' Int. Arch. Allergy, 3, 293.
PARISH, W. E. (1970). 'Homologous serum passivecutaneous anaphylaxis in the guinea pigs mediatedby two yj or yi-type heat-stable globulins and anon-y1 heat labile reagin.'J. Immunol., 105, 1296.
RUSSELL LITTLE, J. and EISEN, H. N. (1967). Methods inImmunology and Immunochemistry (Ed. by C. A. Williamsand M. W. Chase). p. 128. Academic Press, Inc.New York.
SCHEIDEGGER, J. J. (1955). 'Une micromethode del'Immune electrophorese.' Int. Arch. Allergy, 7, 103.
STECHSCHULTE, D., ORANGE, R. and AUSTEN, F. (1970).'Immunochemical and biologic properties of ratIgE. I. Immunochemical identification of rat IgE.'J. Immunol., 105, 1082.
VAN Ess, L., DAN HARINCK, H. and PONDMAN, K. W.(1970). 'The specificity of skin-sensitizing antibodiesin the guinea pig.' Clin. exp. Immunol., 6, 741.
ZVAIFLER, N. and BECKER, E. (1966). 'Rabbit anaphy-lactic antibody.'J. exp. Med., 123, 935.