Embed Size (px)
Citation preview

MECCANISMI DI RIPARAZIONE
DEL DNA

MUTAZIONI SPONTANEE ED INDOTTE
Il danno al DNA non riparato può portare a mutazioni che causano malattie o morte delle cellule.
Le mutazioni derivano da cambiamenti della sequenza dei nucleotidi del DNA, o da delezioni, inserzioni o riarrangiamenti di sequenze di DNA genomico.
Mutazione spontanea si verifica come risultato di processi naturali che avvengono nelle cellule (errori di replicazione).
Mutazione indotta si verifica come risultato di interazioni del DNA con un agente od un mutageno esterno che provoca un danno al DNA.
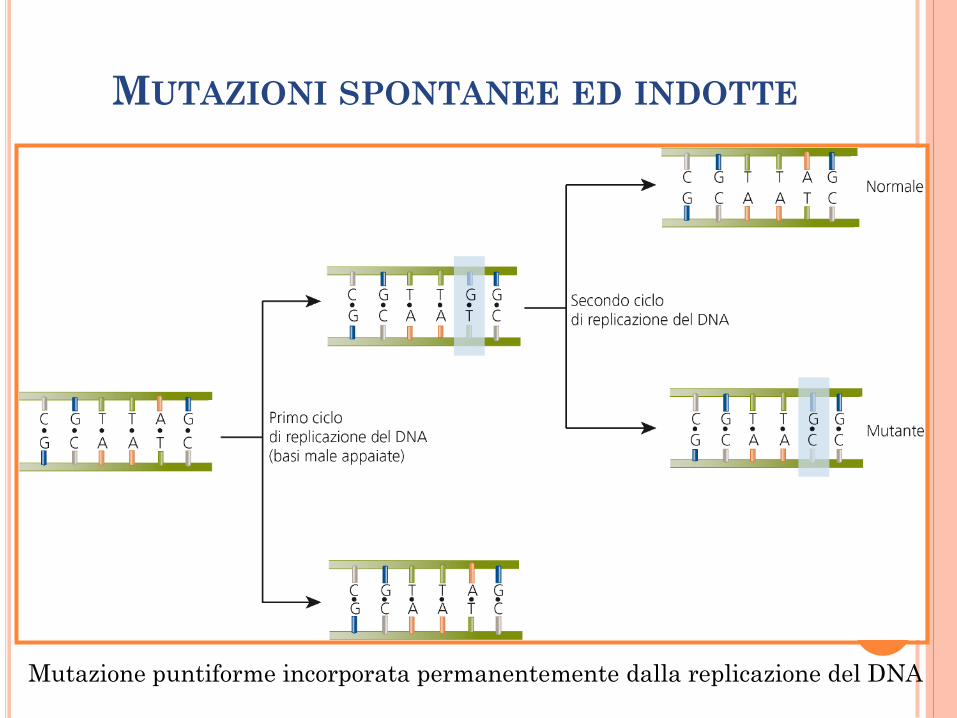
MUTAZIONI SPONTANEE ED INDOTTE
Mutazione puntiforme incorporata permanentemente dalla replicazione del DNA
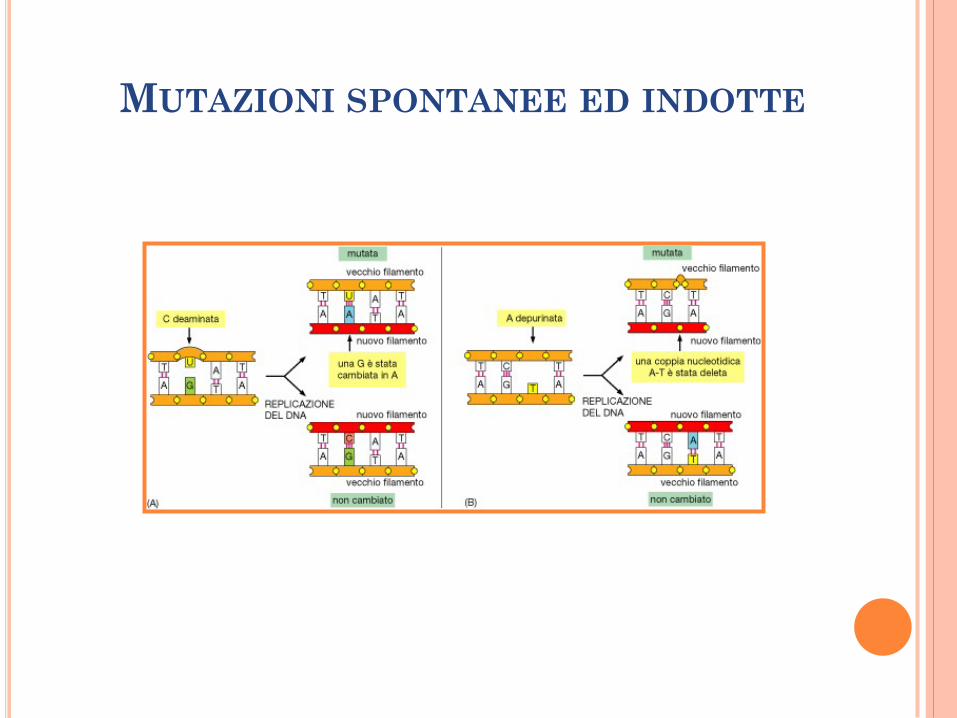
MUTAZIONI SPONTANEE ED INDOTTE

Le mutazioni sono importanti come fonte
principale di variazione genetica cambiamenti
evolutivi.
Le mutazioni possono avere conseguenze deleterie
o (raramente) vantaggiose per un organismo o per i
suoi discendenti.
Gli organismi mutanti sono strumenti importanti
per la caratterizzazione dei geni coinvolti nei
processi cellulari.
Mutazioni puntiformi: mutazioni che alterano
una sola coppia di nucleotidi.
Espansioni delle ripetizioni trinucleotidiche.
Inserzioni o delezioni.
Grandi riarrangiamenti cromosomici.
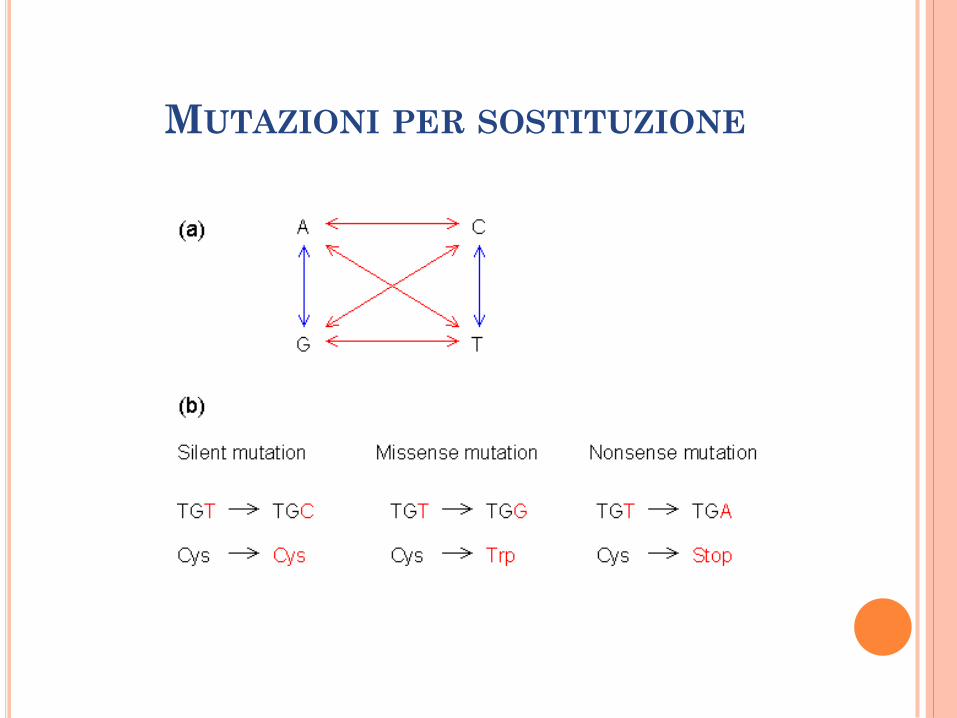
MUTAZIONI PER SOSTITUZIONE

MUTAZIONI PER SOSTITUZIONE
Le sostituzioni nucleotidiche spontanee tendono ad essere transizioni.
Nel genoma umano il rapporto trans:transv è 2:1.
Effetto fenotipico: se alterano un nucleotide cruciale in una regione di regolazione di un gene, nello stampo di una molecola funzionale di RNA o provochino mutazioni in un gene che codifica per una proteina.
Mutazioni nei geni codificanti per proteine:
Silenti;
Missenso (anemia falciforme: AT TA);
Nonsenso (creazione di un nuovo codone di stop).
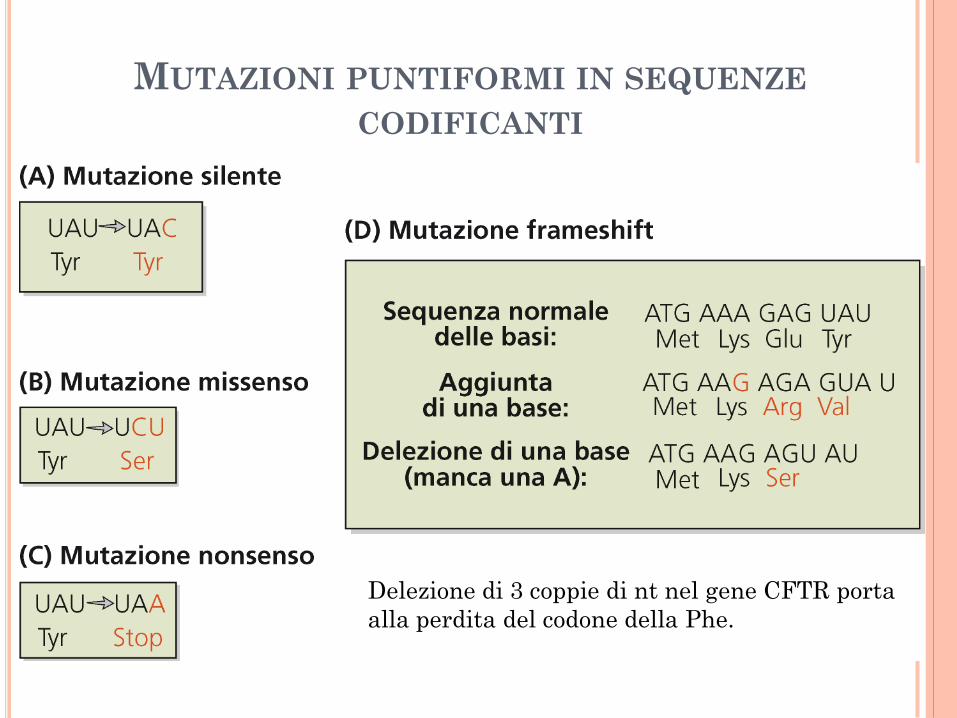
MUTAZIONI PUNTIFORMI IN SEQUENZE
CODIFICANTI
Delezione di 3 coppie di nt nel gene CFTR porta
alla perdita del codone della Phe.
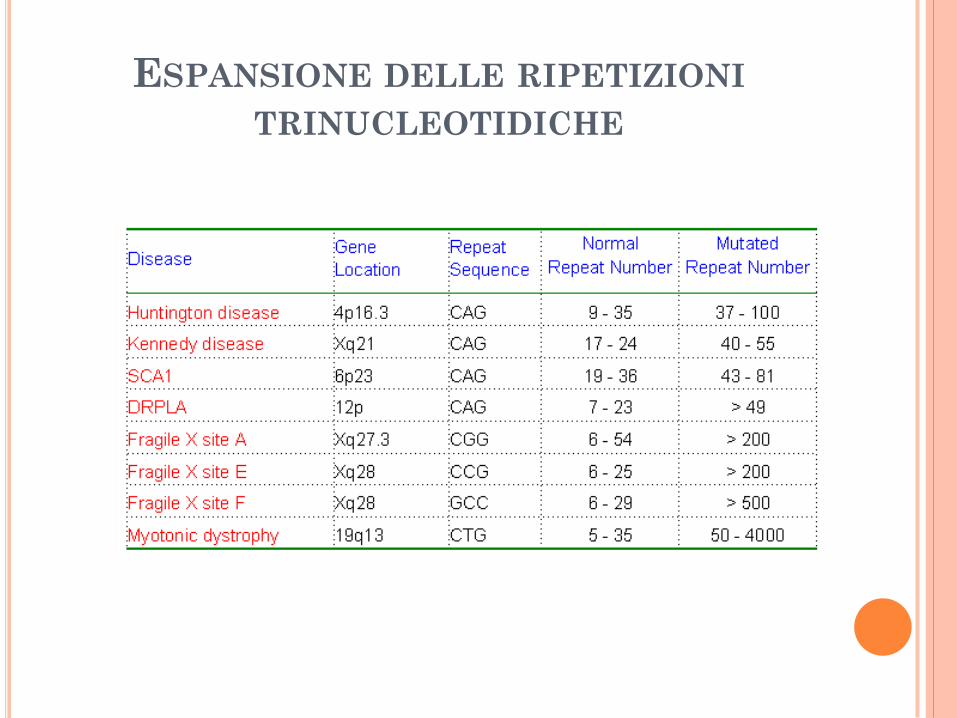
ESPANSIONE DELLE RIPETIZIONI
TRINUCLEOTIDICHE

CLASSI GENERALI DEL DANNO AL DNA
Tre classi principali di danno al DNA:
Cambiamenti di singole basi;
Distorsione strutturale;
Danno all’ossatura del DNA.

MUTAZIONI SPONTANEE ED INDOTTE
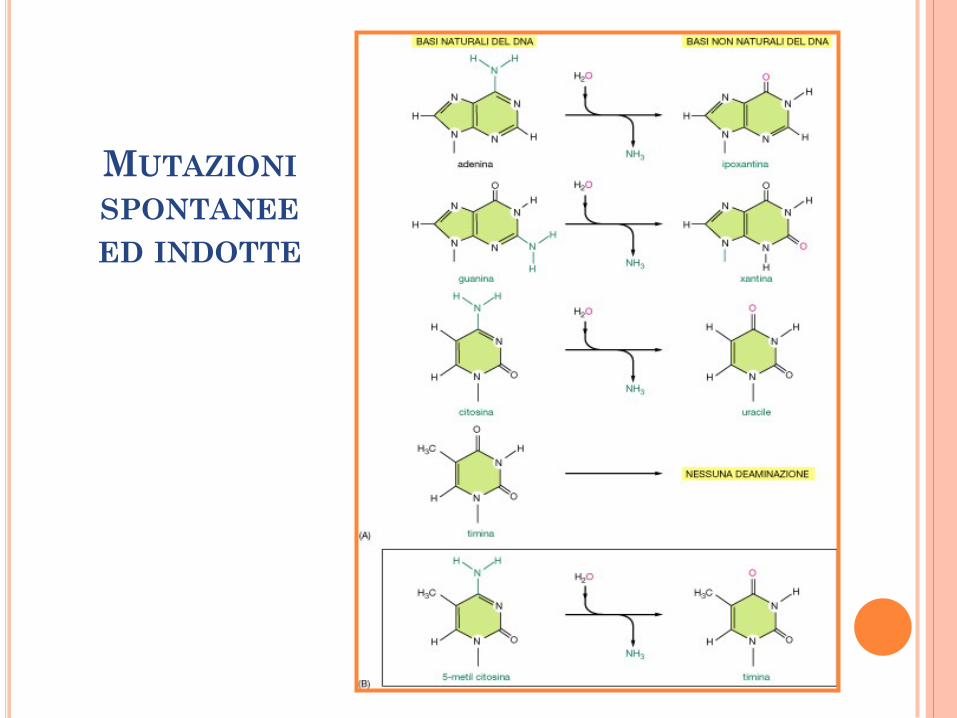
MUTAZIONI
SPONTANEE
ED INDOTTE

CAMBIAMENTI DI SINGOLE BASI
Deaminazione.
Alchilazione per effetto di agenti alchilanti
come le nitrosammine. O6-metilguanina si
accoppia in modo sbagliato con T.
Ossidazione agenti ossidanti (radiazioni
ionizzanti od agenti chimici che generano radicali
liberi) G diventa OxoG che si puo accoppiare
con A.
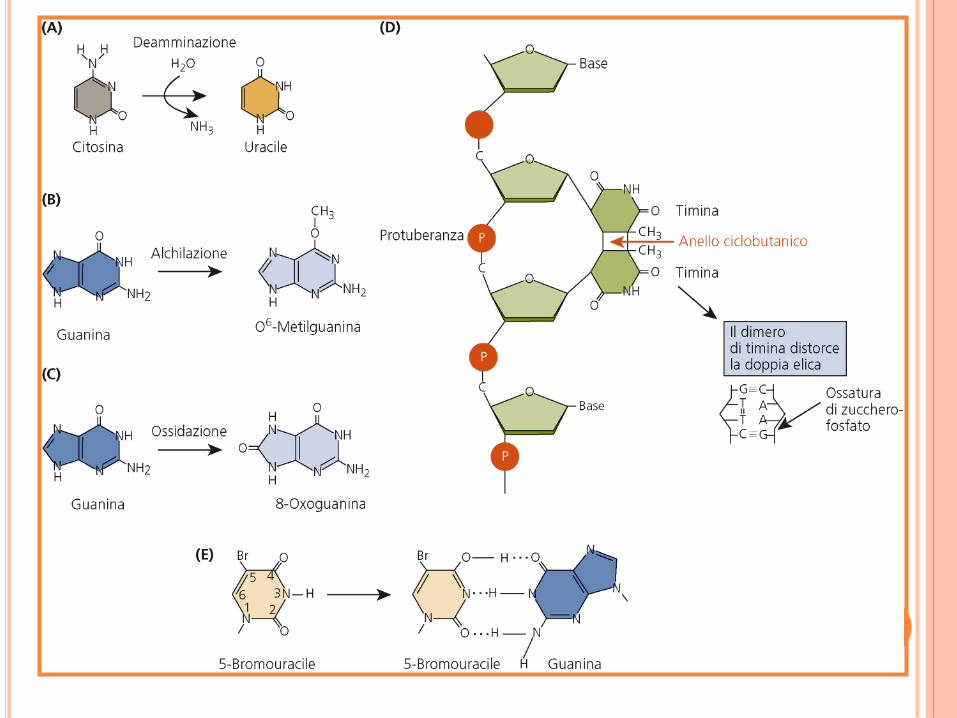
MUTAZIONI SPONTANEE ED INDOTTE
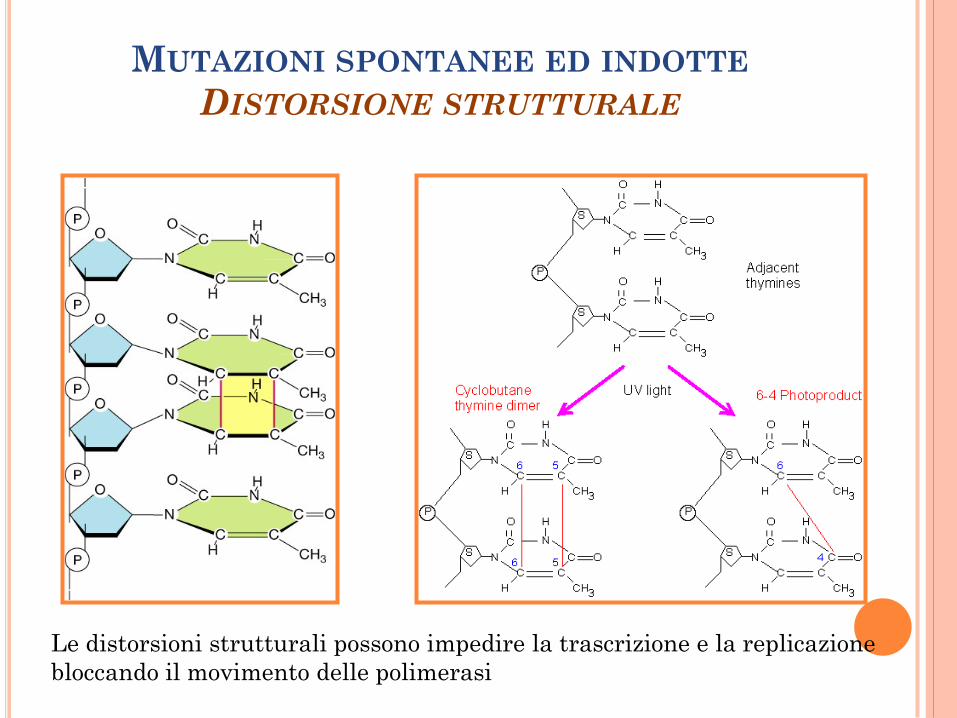
MUTAZIONI SPONTANEE ED INDOTTE
DISTORSIONE STRUTTURALE
Le distorsioni strutturali possono impedire la trascrizione e la replicazione
bloccando il movimento delle polimerasi

Addotti voluminosi possono essere indotti da
mutagenesi chimica.
Agenti intercalanti come il bromuro di etidio
contengono diversi anelli policiclici, così che si
inseriscono facilmente tra le basi del DNA
distorsione della doppia elica del DNA.
Analoghi delle basi azotate, come il 5-
bromouracile, un analogo della timina, che però
si può accoppiare a G.

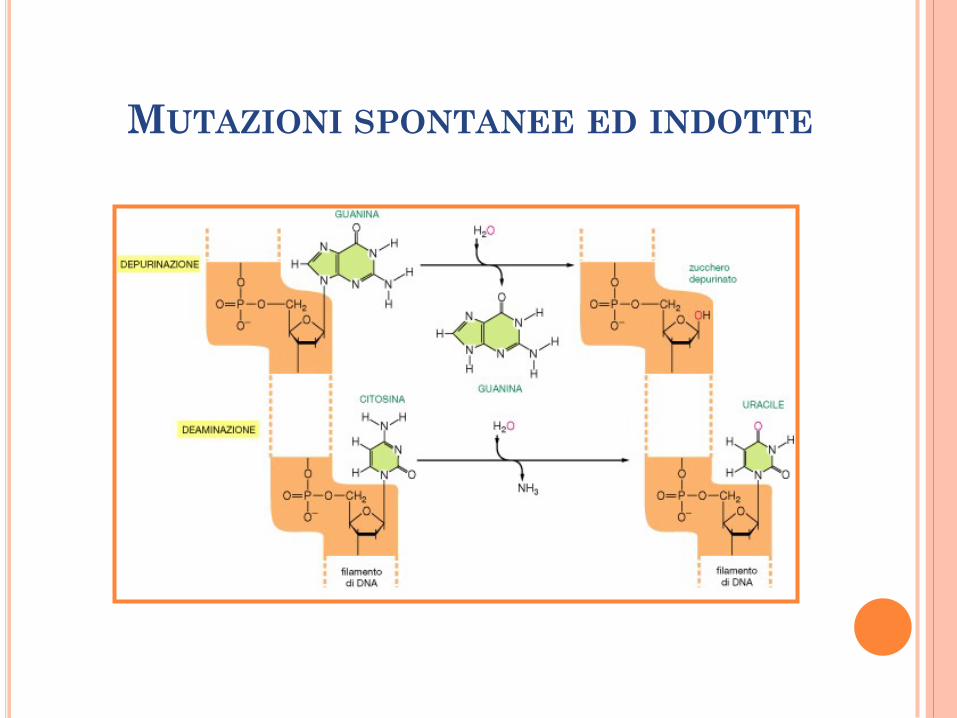
DANNO ALL’OSSATURA DEL DNA
Formazione di siti abasici: idrolisi spontanea del
legame glicosidico tra lo zucchero e la base
azotata (frequente per le purine).
Rottura a doppio filamento indotta da radiazioni
ionizzanti (raggi X, radioisotopi) e da composti
chimici.
Le rotture al doppio filamento sono il tipo più
grave di danno al DNA .

RISPOSTA CELLULARE AL DANNO AL
DNA
Aggiramento del danno.
Inversione del danno.
Rimozione del danno.
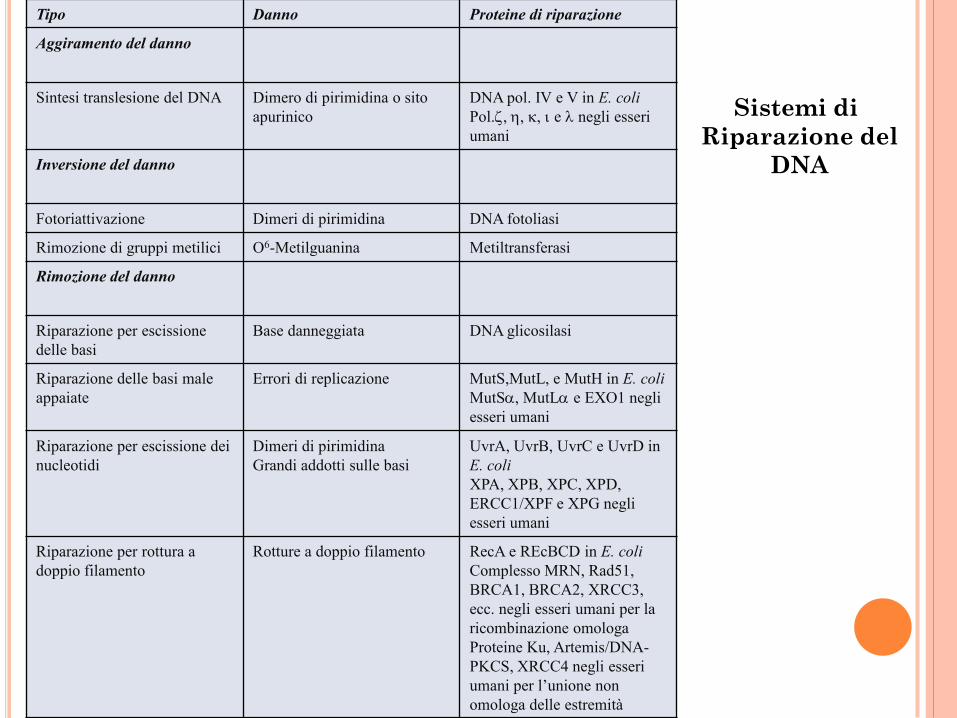
Tipo Danno Proteine di riparazione
Aggiramento del danno
Sintesi translesione del DNA Dimero di pirimidina o sito
apurinico
DNA pol. IV e V in E. coli
Pol., , , e negli esseri
umani
Inversione del danno
Fotoriattivazione Dimeri di pirimidina DNA fotoliasi
Rimozione di gruppi metilici O6-Metilguanina Metiltransferasi
Rimozione del danno
Riparazione per escissione
delle basi
Base danneggiata DNA glicosilasi
Riparazione delle basi male
appaiate
Errori di replicazione MutS,MutL, e MutH in E. coli
MutS, MutL e EXO1 negli
esseri umani
Riparazione per escissione dei
nucleotidi
Dimeri di pirimidina
Grandi addotti sulle basi
UvrA, UvrB, UvrC e UvrD in
E. coli
XPA, XPB, XPC, XPD,
ERCC1/XPF e XPG negli
esseri umani
Riparazione per rottura a
doppio filamento
Rotture a doppio filamento RecA e REcBCD in E. coli
Complesso MRN, Rad51,
BRCA1, BRCA2, XRCC3,
ecc. negli esseri umani per la
ricombinazione omologa
Proteine Ku, Artemis/DNA-
PKCS, XRCC4 negli esseri
umani per l’unione non
omologa delle estremità
Sistemi di
Riparazione del
DNA
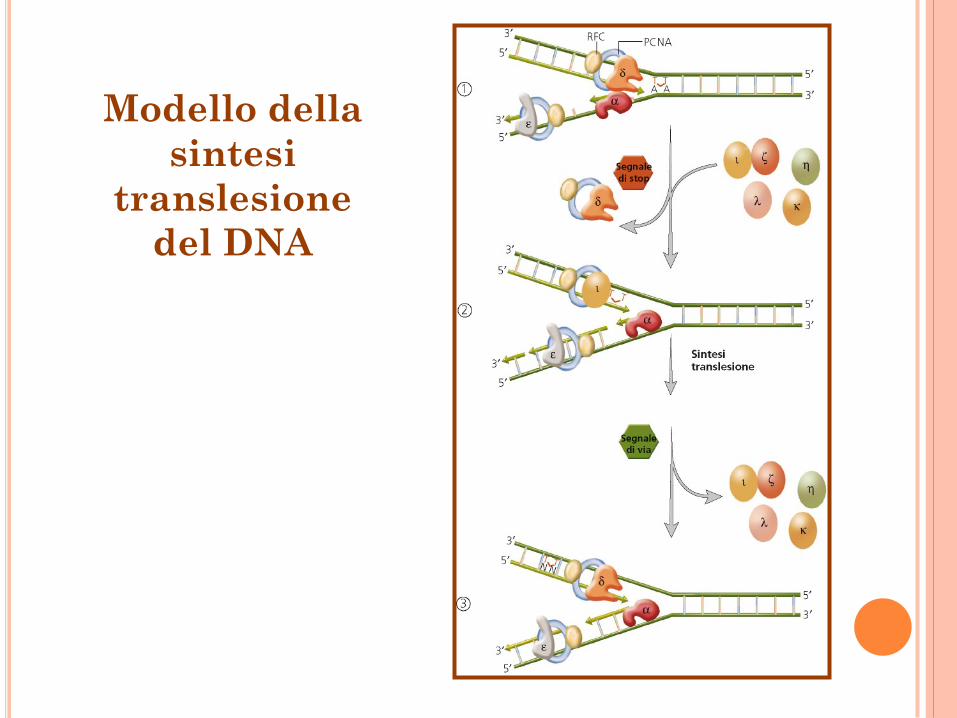
Modello della
sintesi
translesione
del DNA
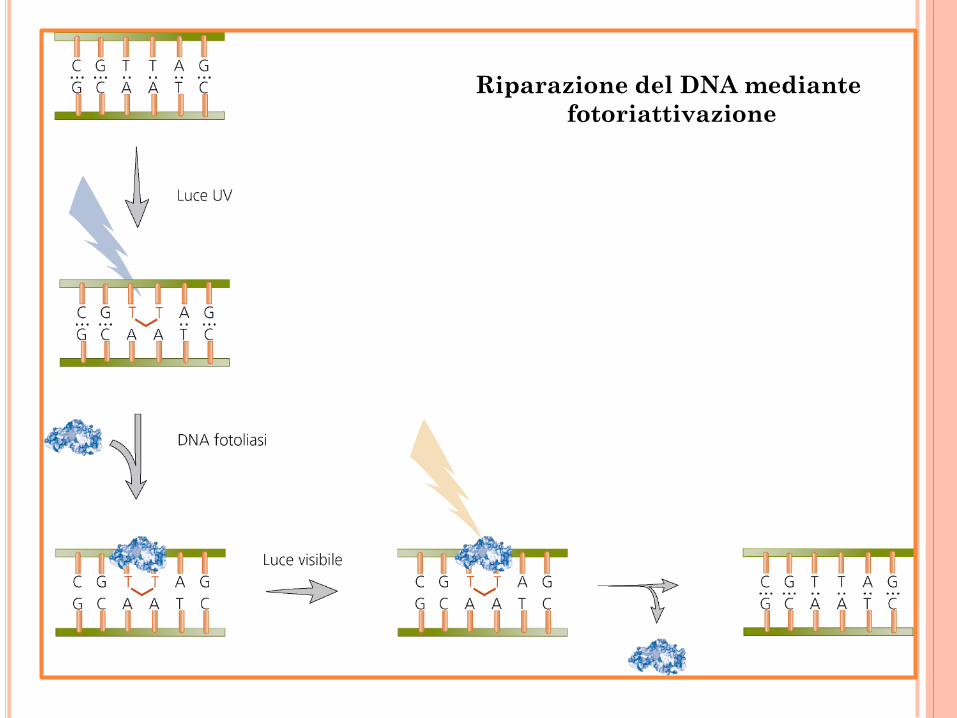
Riparazione del DNA mediante
fotoriattivazione
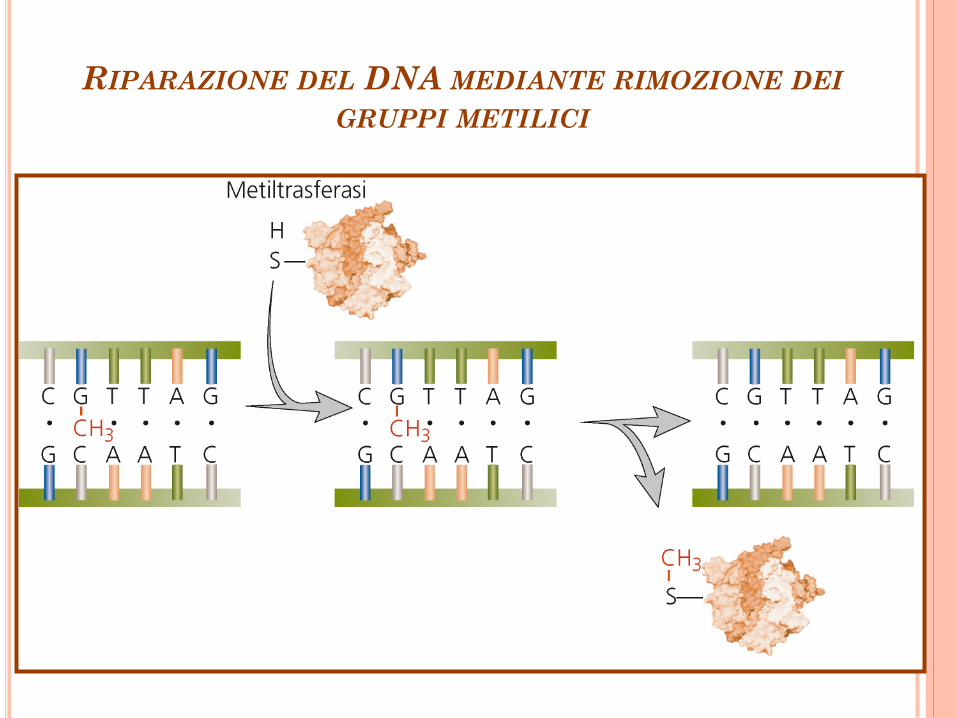
RIPARAZIONE DEL DNA MEDIANTE RIMOZIONE DEI
GRUPPI METILICI
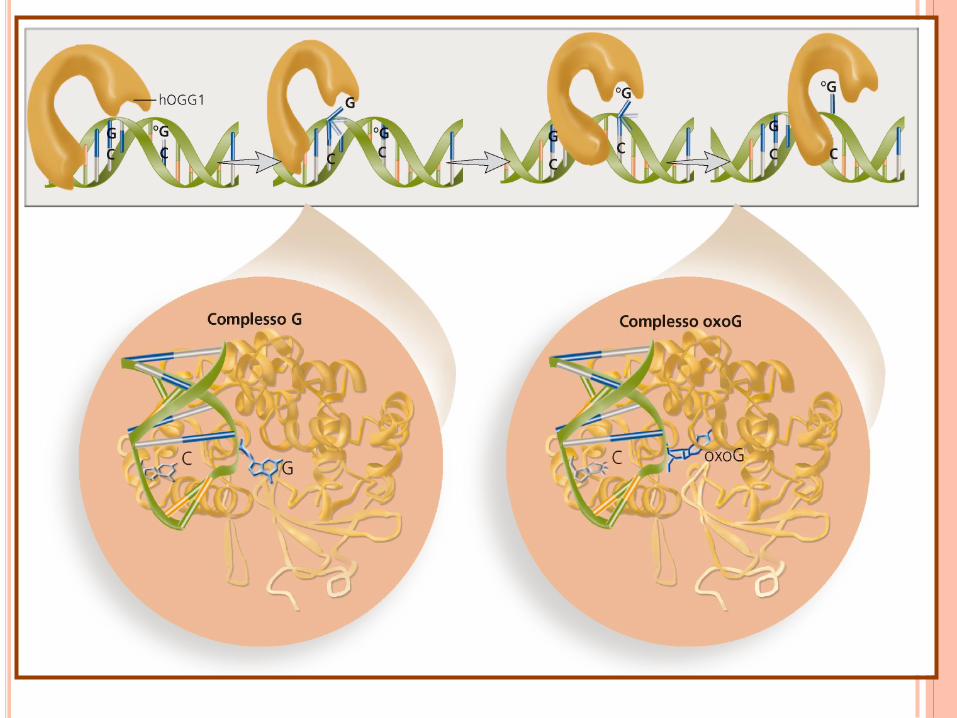
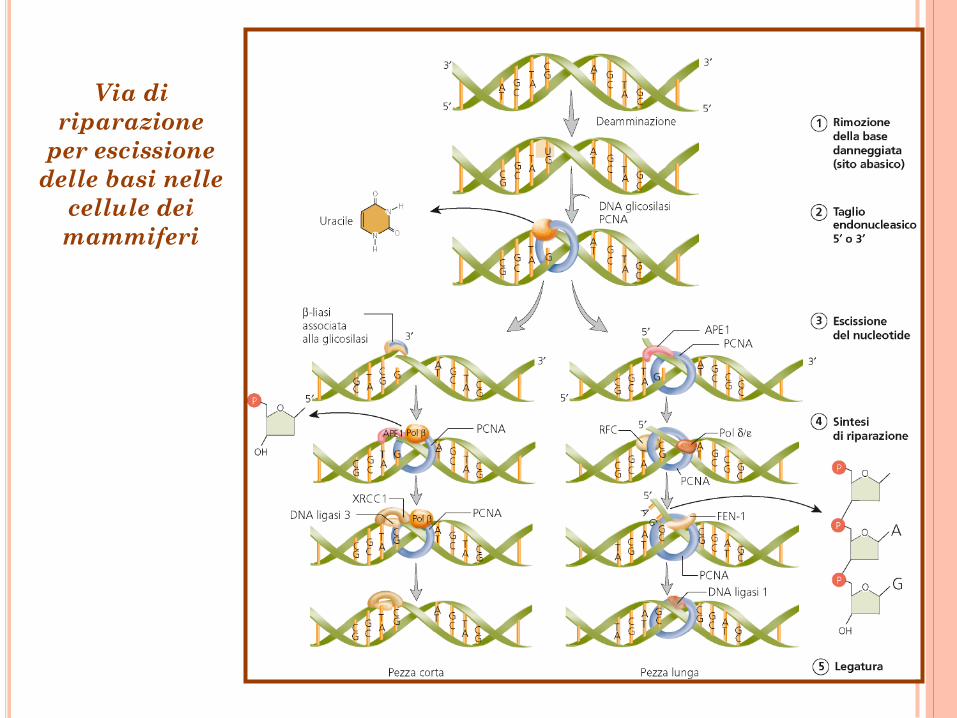
Via di
riparazione
per escissione
delle basi nelle
cellule dei
mammiferi
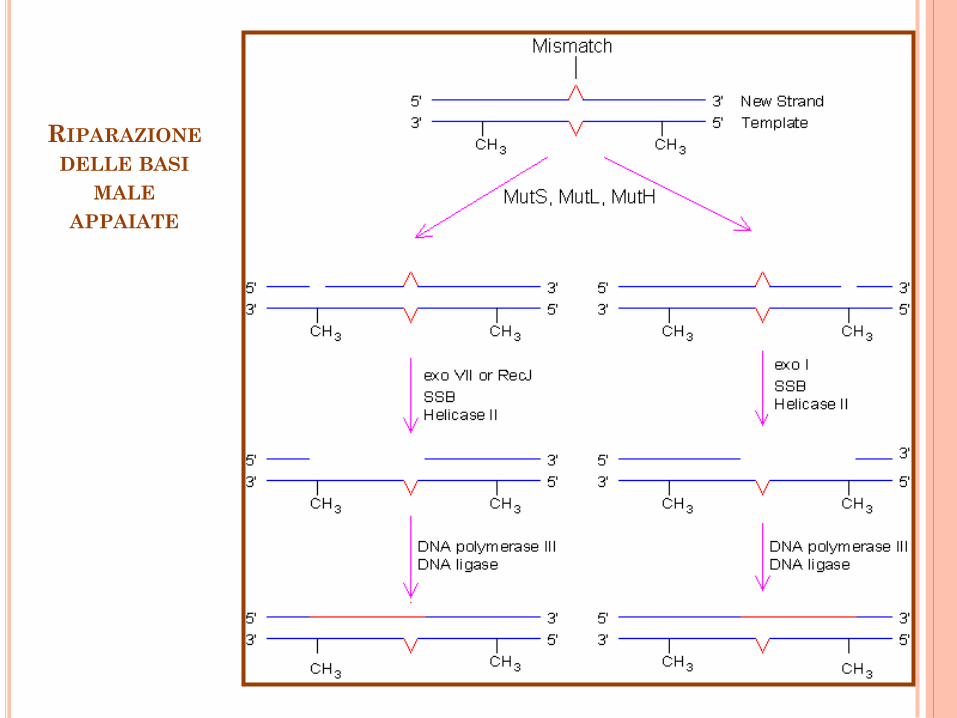
RIPARAZIONE
DELLE BASI
MALE
APPAIATE

Riparazione delle basi male appaiate
La riparazione delle basi male appiate dipende
da numerosi fattori presenti nelle cellule umane,
fra cui MutS (eterodimero) o MutS, MutL
(eterodimero), esonucleasi EXO1, DNA pol. ,
PCNA, caricatore della pinza (RFC), proteina che
lega il singolo filamento (RPA), e proteina
cromosomica non istonica HMGB1.
Il complesso MutS- MutL sul DNA con
appaiamento sbagliato scatena l’attivazione del
macchinario di riparazione.
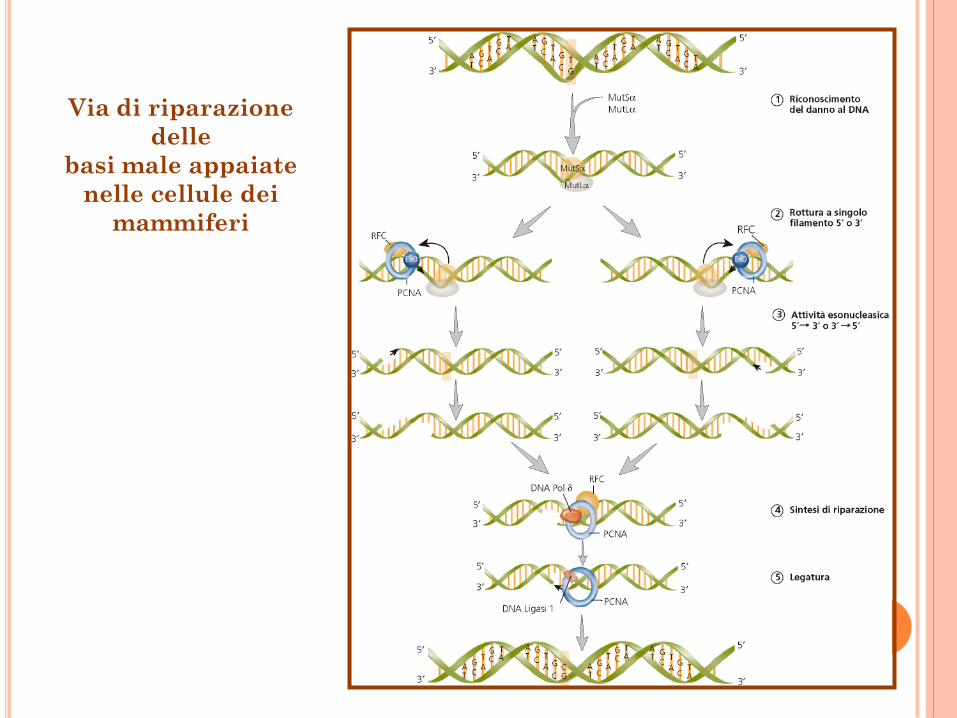
Via di riparazione
delle
basi male appaiate
nelle cellule dei
mammiferi

CANCRO COLON-RETTALE EREDITARIO NON
ASSOCIATO A POLIPOSI
L’80% dei cancri del colon è sporadico, mentre un
20% ha una suscettibilità ereditaria alla malattia.
Il cancro colon-rettale non associato a poliposi
(HPNCC) è la forma ereditaria più comune di cancro
colonrettale.
E’ una condizione autosomica dominante dovuta a
mutazioni in uno dei diversi geni della riparazione
delle basi male appaiate.
Le mutazioni nei geni MutS o MutL assommano a
più del 90% delle mutazioni nelle famiglie con
HPNCC.

Progressione verso l’HPNCC:
1. Mutazione germinale in un allele dei geni di
riparazione delle basi male appaiate.
2. Perdita somatica dell’allele normale.
3. Difetto del meccanismo di riparazione delle
basi male appaiate.
4. Accumulo di errori durante la replicazione del
DNA.
5. Instabilità dei microsatelliti.
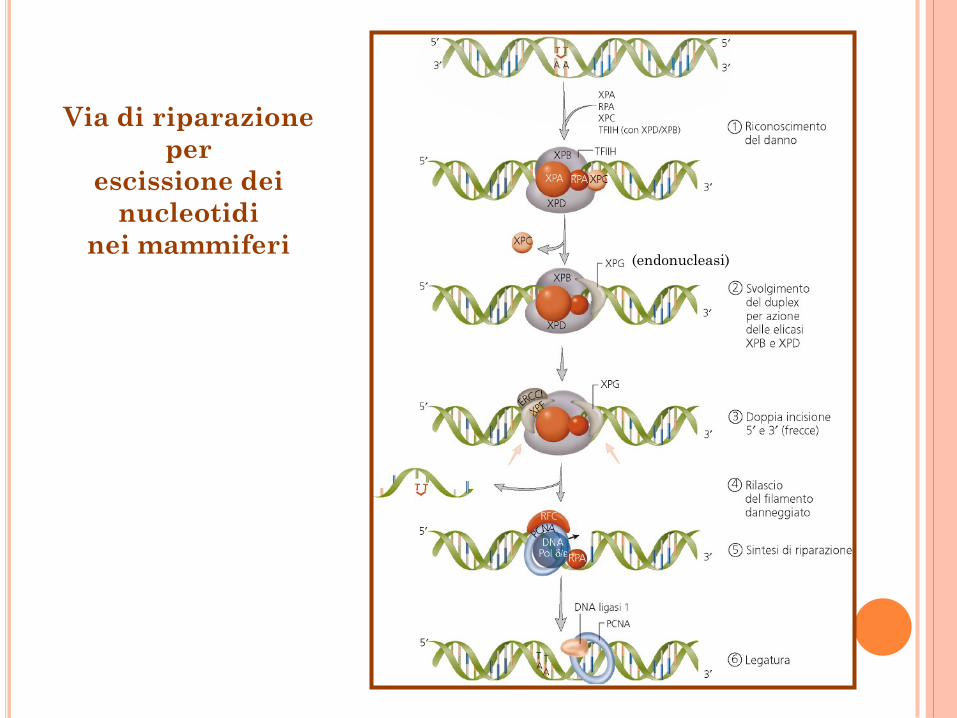
Via di riparazione
per
escissione dei
nucleotidi
nei mammiferi (endonucleasi)
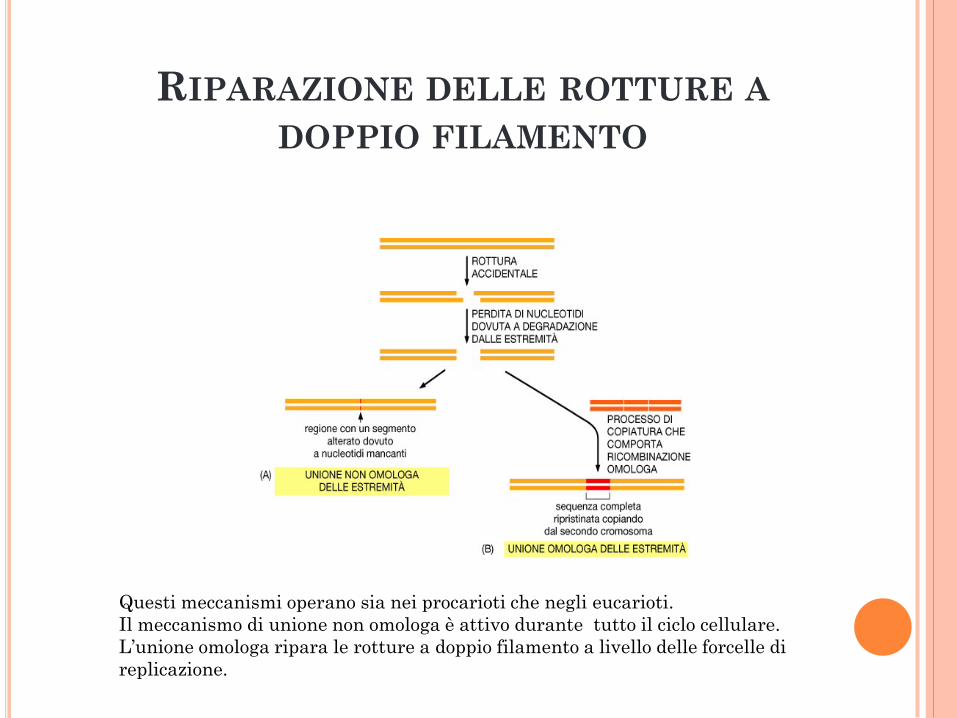
RIPARAZIONE DELLE ROTTURE A
DOPPIO FILAMENTO
Questi meccanismi operano sia nei procarioti che negli eucarioti.
Il meccanismo di unione non omologa è attivo durante tutto il ciclo cellulare.
L’unione omologa ripara le rotture a doppio filamento a livello delle forcelle di
replicazione.
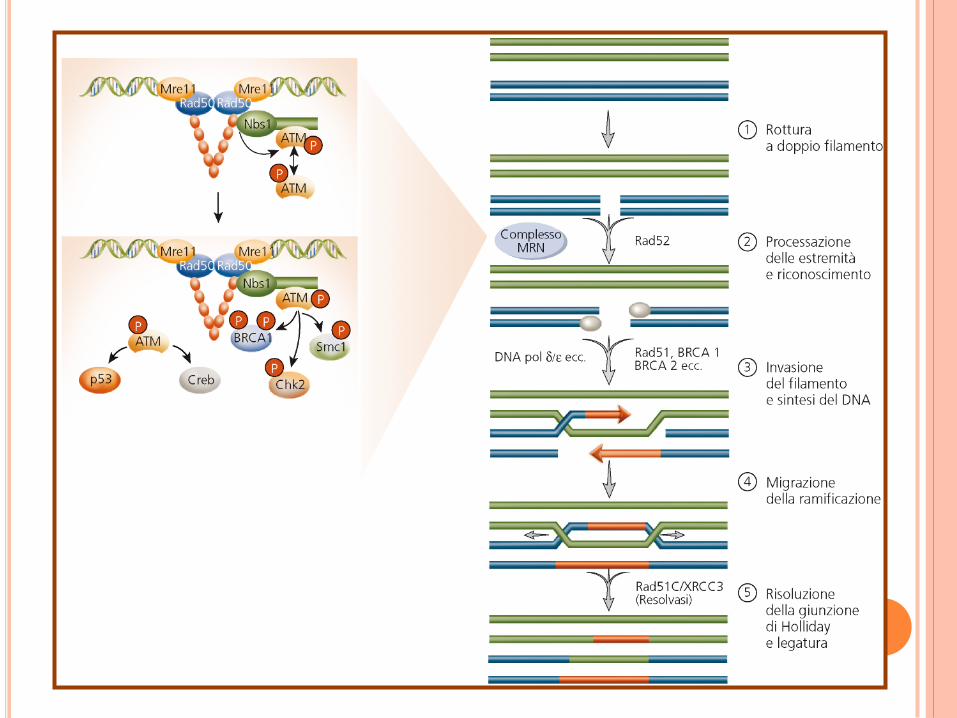

UNIONE OMOLOGA –RICOMBINAZIONE
OMOLOGA
Proteine coinvolte:
Ser/Thr chinasi nucleare, ATM (trasduttore chiave)
aumenta l’attività di questa chin
asi.
ATM fosforila proteine coinvolte nella riparazione del DNA
(BRCA1) e nel controllo del ciclo cellulare (p53; proteina
che sopprime i tumori).
Localizzazione nei siti di rottura di: ATM, proteina Rad52
il complesso Mre11-Rad52-Nbs1 (MRN) inizia la
riparazione (Mre11: esonucleasi 3’ 5’.
Il DNA a singolo filamento sono riconosciuti da: Rad51.
Altre proteine sono coinvoltenella ricombinazione omologa:
Rad54, Rad55, Rad57, BRCA1 e BRCA2.

STRUTTURA DELLA GIUNZIONE DI
HOLLIDAY
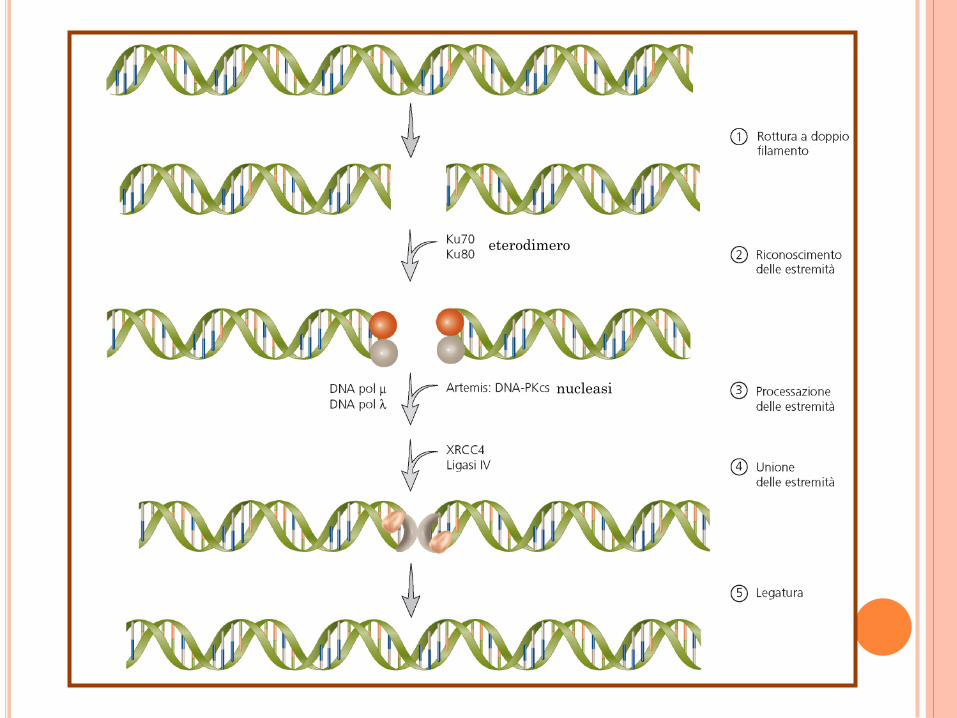
eterodimero
nucleasi





























