Embed Size (px)
Citation preview

i
Metabolic and Ecological Study of Environmental Pentose Utilizing
Bacteria (E-PUB)
By
Farhana Sharmin
BSc (Hons) In Microbiology
Gono University, Bangladesh
Master of Science (Food and Bio Process Technology,
Asian Institute of Technology (AIT), Bangkok, Thailand
A thesis submitted in partial fulfilment of the requirements for the degree of
Doctor of Philosophy
School of Biomedical Sciences
Faculty of Health
Queensland University of Technology
2012

i
TABLE OF CONTENTS
LIST OF TABLES IV
LIST OF FIGURES VI
LIST OF ABBREVIATIONS VIII
ACKNOWLEDGEMENTS XII
ABSTRACT 3
CHAPTER 1 5
INTRODUCTION, HYPOTHESIS, AIMS AND OBJECTIVES 5
1.1 BACKGROUND 5
1.2 RESEARCH PROBLEM AND HYPOTHESIS 7
1.3 AIMS AND OBJECTIVES OF RESEARCH 7
1.4 RESEARCH PLAN 9
CHAPTER 2 11
LITERATURE REVIEW 11
2.0 INTRODUCTION 11
2.1 BRIEF HISTORICAL BACKGROUND 11
2.2 THE CHEMISTRY OF PLANT BIOMASS 13
2.3 MICROBIAL ASPECTS OF FERMENTATION TECHNOLOGY 19
2.4 METHODOLOGY 29
CHAPTER 3
ISOLATION AND IDENTIFICATION OF ENVIRONMENTAL PENTOSE-UTILIZING BACTERIA 37
3.0 SUMMARY 37
3.1 INTRODUCTION 37
3.2 MATERIALS AND METHODS 38
3.3 RESULTS 43
3.4 DISCUSSION 47
3.5 CONCLUSIONS 49
CHAPTER 4
CATABOLIC CHARACTERISTICS OF ENVIRONMENTAL PENTOSE-UTILIZING BACTERIA (E-PUB) 50
4.0 SUMMARY 50
4.1 INTRODUCTION 50
4.2 MATERIALS AND METHODS 52

ii
4.3 RESULTS 53
4.4 DISCUSSION 62
4.5 CONCLUSIONS 68
CHAPTER 5
ANABOLIC CHARACTERISTICS OF ENVIRONMENTAL PENTOSE-UTILIZING BACTERIA 69
5.0 SUMMARY 69
5.1 INTRODUCTION 69
5.2 MATERIALS AND METHODS 71
5.3 RESULTS 74
5.4 DISCUSSION 81
5.5 CONCLUSIONS 87
CHAPTER 6
ECOLOGICAL STUDY OF ENVIRONMENTAL PENTOSE-UTILIZING BACTERIA BY DENATURING
GRADIENT GEL ELECTROPHORESIS (DGGE) 88
6.0 SUMMARY 88
6.1 INTRODUCTION 88
6.2 MATERIALS AND METHODS 90
6.3 RESULTS 94
6.4 DISCUSSION 103
6.5 CONCLUSIONS 109
CHAPTER 7
TAXONOMIC ANALYSIS OF PENTOSE-RICH NATURAL ENVIRONMENTS USING A HIGH DENSITY
OLIGONUCLEOTIDE MICROARRAY (PHYLOCHIP) TECHNOLOGY 111
7.0 SUMMARY 111
7.1 INTRODUCTION 111
7.2 MATERIALS AND METHODS 112
7.3 RESULTS 115
7.4 DISCUSSION 123
7.5 CONCLUSIONS 126
CHAPTER 8 128
SUMMARY, CONCLUSION AND FUTURE WORK 128
8.1 REVISITING THE HYPOTHESIS AND AIMS 128
8.2 SUMMARY OF FINDINGS 129

iii
8.3 SIGNIFICANCE OF FINDINGS 132
8.4 FUTURE DIRECTIONS 133
REFERENCES 137
APPENDIX A ERROR! BOOKMARK NOT DEFINED.
APPENDIX B ERROR! BOOKMARK NOT DEFINED.

iv
LIST OF TABLES
Table 2.1: Comparison of various lignocellulosic raw materials...................................................17
Table 2.2: List of pentose utilizing microorganisms (adapted from Singh and Mishra, 1995).......23
Table 2.3: List of microbial products from pentose fermentation (Singh and Mishra, 1995).......27
Table 3.1: Location of Collected Samples.....................................................................................39
Table 3.2: PCR reaction mixture details........................................................................................42
Table 3.3: Characteristics of selected isolates from Maryborough (A)..........................................43
Table 3.4: Characteristics of selected isolates from Maryborough (B)......................................... 44
Table 3.5: Characteristics of selected isolates from Proserpine....................................................44
Table 3.6: 16S rDNA sequencing results of unknown isolates......................................................46
Table 4.1: Specific growth rates of E-PUB cultures.......................................................................56
Table 4.2: Statistical analysis of p values of specific growth rates................................................57
Table 4.3: Analysis of diauxie growth............................................................................................59
Table 4.4: P value of significance specific growth rate of combination of glucose and pentose
sugars............................................................................................................................................60
Table 4.5: Amount of glucose at the second lag phase of the growth medium............................61
Table 4.6: Ratio of specific growth rate (µ) for xylose utilization as a single sugar compared to
that of the same sugar as part of a dual carbon source system.....................................................61
Table 5.1: HPLC injector programme for amino acid detection.....................................................73
Table 5.2: Names of amino acids detected and their elution times..............................................74
Table 5.3: Amino acids produced by E-PUB isolates …………………………….…….…………………………….75
Table 5.4: Performance characteristics of HPLC UV detection of amino acid...............................77
Table 5.5: Amino acid production using single and dual carbon sources as substrates………..……80
Table 5.6: Total biomass measured from single pentose and dual sugar carbon substrate.........80
Table 5.7: Yield of amino acid expressed in mg per g of biomass .................................................81
Table 6.1: List of the samples collected from different sugar mills...............................................90
Table 6.2: 16S rDNA sequencing results including samples and DGGE band details...................102
Table 6.3: 16S rDNA sequencing result with closest matched Genus/species based on the
percentage (%) of similarity.........................................................................................................103

v
Table 7.1: Richness of the bacterial taxa present in environmental samples from sugar-cane
processing sites............................................................................................................................116
Table 7.2: Evenness (Shannon’s diversity index; H’) of bacterial communities present from
sugar-cane processing sites.........................................................................................................116
Table 7.3: Summary of CAP data showing the significance of sample types and location effects
on bacterial community composition..........................................................................................121
Table 7.4: Summary SIMPER data...............................................................................................123

vi
LIST OF FIGURES
Figure 2.1: Chemical structure of starch..................................................................................15
Figure 2.2: Chemical structure of pectin.......................................................................................15
Figure 2.3: Structure of lignocellulose...........................................................................................16
Figure 2.4: Chemical structure of xylan and xylose formation ....................................................19
Figure 2.5: o-phthalaldehyde (OPA) and mercaptoethanol reaction...........................................32
Figure 3.1: Map of the Queensland sugar mill regions.................................................................38
Figure 3.2: 16S rDNA PCR amplification image after gel electrophoresis……….......................….45
Figure 4.1 (a-g): Comparison of growth curves for various carbon sources..................................55
Figure 4.2: Results of the growth measurements for the various cultures...................................58
Figure 4.3: Metabolic pathway of breakdown pentose and glucose sugars….....................…….63
Figure 4.4: Metabolic pathways for the breakdown of sugars (xylose included.......................64
Figure 5.1: Chromatogram of a mixture of 0.5 µL amino acids standards…………………..............…74
Figure 5.2: Chromatograms of the amino acid mixture present in microbial
cultures..................776
Figure 5.3: HPLC for LB medium-blank analysis.............................................................................78
Figure 5.4: End-products of dual-sugar metabolism (glucose and pentose)…….......................... 79
Figure 5.5: Formation of intracellular and extracellular amino acids...........................................82
Figure 5.6: Glycine biosynthesis…………………………………….……………………………………………………….. 86
Figure 6.1: Diagram showing a typical sugar processing mill. .........…………………………………………..91
Figure 6.2: Community fingerprint analysis using the Bray-Curtis method................................96
Figure 6.3: DGGE profiles of bacterial community structures in liquid samples..........................97
Figure 6.4: Phylogenetic tree analysis of liquid samples by.........................................................97
Figure 6.5: Non-metric multi-dimensional scaling (MDS) plot analysis of DGGE bands .............99
Figure 6.6: DGGE profiles of bacterial community structures in soil samples.............................100
Figure 6.7: Phylogenetic tree analysis of solid samples...............................................................100
Figure 6.8: DGGE gel image indicating the lane numbers............................................................101
Figure 7.1: Dominant bacterial phyla present. ...........................................................................117

vii
Figure 7.2: Distribution of bacterial Classes present within the four dominant Phyla ...............118
Figure 7.3: Heat map / 2-way clustering of samples according to PhyloChip OTU’s. Clustering
based on Euclidean distance using complete linkage method.....................................................120
Figure 7.4: nMDS ordination plot showing similarity in bacterial community structure.............122

viii
LIST of ABBREVIATIONS
A. Actinomyces
Ala Alanine
AT Annealing Temperature
Arg Arginine
Asp Aspartic acid
bp Base Pairs
BLAST Basic Local Alignment Search Tool
C. Corynebacterium
CAP Canonical Analysis of Principal
cDNA Copy DNA
Cys Cysteine
CV Coefficient of Variation
DGGE Denaturing Gradient Gel Electrophoresis
DNA Deoxyribonucleic Acid
dNTPs Nucleoside Triphosphates
dsDNA Double Stranded DNA
E Shannon Evenness Index
EDTA Ethylenediaminetetracetic acid
FISH Fluorescence In Situ Hydridization
FMOC Fluorenylmethoxycarbonyl
GC Gas Chromatography
gDNA Genomic DNA
Glu Glutamic acid
Gly Glycine
g Gram (weight)
H Shannon-Wiener diversity index
h Hour
HPLC High Performance Liquid Chromatography
His Histidine

ix
Ile Isoleucine
LB Luria-Bertani
lys Lysine
µL Microlitre
µM/ µmol Micromolar
M Molar
mL Millilitre
min Minute
mMol milli-Mole
MDS Multidimensional scaling
N. Nocardia
ng Nano gram
OPA o-Phthaldialdehyde
OTU Operational Taxonomic Unit
OD Optical Density
phe Phenylalanine
% Percentage
PCR Polymerase Chain Reaction
P. Propionibacterium
pmol Picomole
qPCR Quantitative real-time PCR
RE Restriction Endonuclease
R. Rhodococcus
RNA Ribonucleic Acid
rRNA Ribosomal RNA
rpm Rotation per Minute
RT-PCR Reverse Transcription PCR
ser Serine
SD Standard Deviation
SNP Single Nucleotide Polymorphisom

x
SSU Small Subunit
TAE Tris Acetate EDTA
TE Tris EDTA
thr Threonine
tyr Tyrosine
U Units
UV Ultraviolet
UPGMA Unweighted Pair Group Arithmetic Mean
W/V Weight/ Volume
List of Supplementary Materials

xi
Appendix A: Thesis-associated Poster/paper presentations
Posters presented at National/International Conferences
1. Farhana Sharmin, John Bartley, Flavia Huygens, Megan Hargreaves (2009). Pentose
Utilizing Corynebacterium sp. Producing Industrial Valuable Amino Acids: 3rd Congress
of European Microbiologists, FEMS, Gothenburg, Sweden and also presented in the
same year at Australian Society for Microbiology conference held in Perth.
2. Farhana Sharmin, Megan Hargreaves (2009), Microbial Metabolism of Pentoses
Released from Lignocellulosic Biomass: 3rd Congress of European Microbiologists,
FEMS; Gothenburg, Sweden.
3. Farhana Sharmin, John Bartley, Flavia Huygens, Megan Hargreaves (2010). PCR-DGGE
fingerprinting for detection of microbial diversity from Queensland sugar industrial
waste. Joint meeting of the NZ Microbiological Society and NZ Society for Biochemistry
& Molecular Biology, University of Auckland, New Zealand.
Papers submitted for Publication
1. Farhana Sharmin, Flavia Huygens, Steve Wakelin and Megan Hargreaves “Firmicutes
dominated the bacterial taxa with in sugar-cane processing plants". Scientific Reports
(under review).
2. Farhana Sharmin, Flavia Huygens and Megan Hargreaves “A review of pentose utilizing
bacteria from bagasse hemicelluolse”. Biotechnology Letter (Manuscript submitted).
3. Farhana Sharmin, Flavia Huygens and Megan Hargraves. “Isolation and Identification and
Ecological Analysis of Environmental Pentose-Utilizing Microbial Community”, Journal of
Applied Microbiology (Manuscript submitted)
4. Farhana Sharmin, Flavia Huygens and Megan Hargreaves. “Study of Catabolic and
Anabolic Characteristics of Environmental Pentose-Utilizing Bacteria”. In Progress
Appendix B: Results and Calculations

xii
ACKNOWLEDGEMENTS
I wish to express a great dept of gratitude to my patient and persistent PhD supervisor, Associate
Professor Megan Hargreaves for sharing her knowledge and for her direction, guidance,
feedback, advice and enormous inspiration during my PhD journey and pregnancy. My thanks
also go to Dr. Flavia Huygens for being such a wonderful and friendly associate supervisor and
for being such a dedicated and thorough research scientist. You have been very generous with
your time during my PhD. Many thanks also to Dr. John Bartley for helping me in the area of
chemistry, especially in the field of HPLC set up. This thesis definitely owes a lot to your insight
and supervision and I hope that it meets your high standards.
I would like to thank Dr Mark Dawson, as an external supervisor and review committee member.
Thank you for being so supportive and thorough with your feedback.
I’d like to acknowledge the members of the EMRG lab and others, who have been an absolute
pleasure to work with, and who have been extremely helpful with their sharing of ideas, skills
and lab time. They include: Irani Rathnayake, Maxim Sheludchenko, Chaminda Ranasinghe,
Phillipa Perrott, Sue Gill, Vincent Chand and Chris Carvalho.
To Charles Wan - thanks for your support in the initial settling in period in the lab, and also for
helping me with molecular tasks, for example primer design, PCR and gel electrophoresis etc. I
would like also to thank Prof Margaret Britz for giving me an opportunity of embarking on this
PhD by approving the scholarship; sharing her immense knowledge and giving me initial
direction and advice during my early stages of this PhD.
My thanks also to the corresponding people of Maryborough, Proserpine and Mackay sugar
Industries for providing samples. I also would like to thank to Dr. James Smith for collecting the
initial samples. I would like to thank Dr. Steven Walkelin for helping me with my Metagenomic
PhyloChip analysis.
I would finally like to acknowledge my family and friends Mum, Dad, sister Rehnuma, brother
Mehedi and husband Jafar who have provided me with much support and encouragement
during my candidature.

xiii
STATEMENT of AUTHORSHIP
The work contained in this thesis has not been previously submitted to meet requirements for an
award at this or any other higher education institution. To the best of my knowledge and belief, the
thesis contains no material previously published or written by another person except where due
reference is made.
S1gnature
�D. 0 7- . 'J.p/2__ Date
Dedication
I dedicate my dissertation to my wonderful family for supporting me and encouraging me through
this whole process. Special thanks are due to Mohammad Amanullah, Mrs. Delowara Ahmed, and
Abu Jafar Siddiquee. My dad and husband kept me grounded through this process, my daughter
Junainah Siddiquee kept me laughing and on my feet, and my parents sacrificed many luxuries
during my upbringing so that I could always have an excellent education.
QUT Verified Signature

Chapter 1 Introduction
3
ABSTRACT Lignocellulosic materials, such as sugar cane bagasse, a waste product of the sugarcane
processing industry, agricultural residues and herbaceous crops, may serve as an abundant and
comparatively cheap feedstock for largescale industrial fermentation, resulting in the production
of marketable end-products. However, the complex structure of lignocellulosic materials, the
presence of various hexose and pentose sugars in the hemicellulose component, and the
presence of various compounds that inhibit the organisms selected for the fermentation
process, all constitute barriers that add to the production costs and make full scale industrial
production economically less feasible. The work presented in this thesis was conducted in order
to screen microorganisms for ability to utilize pentose sugars derived from the sugar mill
industrial waste. A large number of individual bacterial strains were investigated from hemi-
cellulose rich material collected at the Proserpine and Maryborough sugar mills, notably soil
samples from the mill sites. The research conducted to isolation of six pentose-capable Gram-
positive organisms from the actinomycetes group by using pentose as a sole carbon source in
the cultivation process.
The isolates were identified as Corynebacterium glutamicum, Actinomyces odontolyticus,
Nocardia elegans, and Propionibacterium freudenreichii all of which were isolated from the
hemicellulose-enriched soil. Pentose degrading microbes are very rare in the environment, so
this was a significant discovery. Previous research indicated that microbes could degrade
pentose after genetic modification but the microbes discovered in this research were able to
naturally utilize pentose.
Six isolates, identified as four different genera, were investigated for their ability to utilize single
sugars as substrates (glucose, xylose, arabinose or ribose), and also dual sugars as substrates (a
hexose plus a pentose). The results demonstrated that C. glutamicum, A. odontolyticus, N.
elegans, and P. freudenreichii were pentose-capable (able to grow using xylose or other pentose
sugar), and also showed diauxie growth characteristics during the dual-sugar (glucose, in
combination with xylose, arabinose or ribose) carbon source tests. In addition, it was shown
that the isolates displayed very small differences in growth rates when grown on dual sugars as
compared to single sugars, whether pentose or hexose in nature.
The anabolic characteristics of C. glutamicum, A. odontolyticus, N. elegans and P. freudenreichii
were subsequently investigated by qualitative analysis of their end-products, using high
performance liquid chromatography (HPLC). All of the organisms produced arginine and cysteine
after utilization of the pentose substrates alone. In addition, P. freudenreichii produced alanine

Chapter 1 Introduction
4
and glycine. The end-product profile arising from culture with dual carbon sources was also
tested. Interestingly, this time the product was different. All of them produced the amino acid
glycine, when grown on a combination substrate-mix of glucose with xylose, and also glucose
with arabinose. Only N. elegans was able to break down ribose, either singly or in combination
with glucose, and the end-product of metabolism of the glucose plus ribose substrate
combination was glutamic acid.
The ecological analysis of microbial abundance in sugar mill waste was performed using
denaturing gradient gel electrophoresis (DGGE) and also the metagenomic microarray PhyloChip
method. Eleven solid samples and seven liquid samples were investigated. A very complex
bacterial ecosystem was demonstrated in the seven liquid samples after testing with the
PhyloChip method. It was also shown that bagasse leachate was the most different, compared to
all of the other samples, by virtue of its richness in variety of taxa and the complexity of its
bacterial community. The bacterial community in solid samples from Proserpine, Mackay and
Maryborough sugar mills showed huge diversity. The information found from 16S rDNA
sequencing results was that the bacterial genera Brevibacillus, Rhodospirillaceae, Bacillus, Vibrio
and Pseudomonas were present in greatest abundance. In addition, Corynebacterium was also
found in the soil samples.
The metagenomic studies of the sugar mill samples demonstrate two important outcomes:
firstly that the bagasse leachate, as potentially the most pentose-rich sample tested, had the
most complex and diverse bacterial community; and secondly that the pentose-capable isolates
that were initially discovered at the beginning of this study, were not amongst the most
abundant taxonomic groups discovered in the sugar mill samples, and in fact were, as suspected,
very rare. As a bioprospecting exercise, therefore, the study has discovered organisms that are
naturally present, but in very small numbers, in the appropriate natural environment. This has
implications for the industrial application of E-PUB, in that a seeding process using a starter
culture will be necessary for industrial purposes, rather than simply assuming that natural
fermentation might occur.

Chapter 1 Introduction
5
CHAPTER 1
Introduction, Hypothesis, Aims and Objectives
1.1 Background
Microbial utilization of lignocellulosic biomass for the production of commercially valuable
products such as chemicals, liquid fuels, protein enriched food/feed, and preparation of
cellulose polymers, is an attractive approach to help meet energy and food demands of
developed and developing countries. There has been considerable interest in the conversion of
renewable resources into fuels and other products over past years, as petroleum prices rise and
the potential impact of global warming demands technologies that close the carbon cycle
(Govindaswamy and Vane, 2007). The technologies for producing ethanol from plant biomass
constituents such as starch have been available for a number of years. However, the competing
use of starch for food production has focused research on alternative sources, such as the use of
lignocellulosic fractions of plants for production of fuel and chemicals. Government policy in the
USA has mandated that biofuels must increasingly come from sustainable resources, so that
crop wastes in the future will become the primary source of biofuels and other similar products,
rather than carbohydrates such as starch which can be better employed as food products (U.S.
Department of Energy, 2005).
Lignocellulosic biomass contains several polymeric components such as lignin, cellulose and
hemicellulose. Fractionation and enzymatic treatment can therefore yield various product
streams that are rich in phenolics from lignin, glucose from cellulose, and pentoses (mainly
xylose and arabinose) from hemicellulose. Unfortunately, fermentation involving mixtures of
sugars (glucose, xylose, arabinose and others) such as are present in the lignocellulosic biomass,
usually results in the preferential use of glucose due to catabolite repression and consequent
failure to fully utilize all of the available sugars. Commercial fermentation systems aim to
maximize productivity, so that high product yields per unit of microbial biomass are produced,
and the fullest possible use of the carbohydrate source provided, is always the preferred
outcome.
Utilization of the fraction containing hemicellulose in lignocellulosic biomass, is an important
factor in optimizing the economics of biomass-related commercial processes given that

Chapter 1 Introduction
6
hemicellulose makes up a significant proportion (44% carbon content) of the potentially
available carbon for use in fermentation or chemical extraction processes (Hoch, 2007).
Although there are several potential uses for fractions enriched with hemicellulose, such as non-
biological processes for the production of chemicals, the use of hemicellulose carbon as a
fermentable substrate remains an attractive route for producing chemicals that have intrinsic
commercial value. Examples include the production of acids that can be used in the food
industry (such as variety of amino acids, acetic, propionic and lactic acid) or polymer production
(butyrate and lactic acid).
The bioconversion of pentoses derived from hemicellulose remains a bottleneck in development
of industrial fermentation processes, as microbes such as Saccharomyces cerevisiae, that are
known to be able to produce high yields of product, are not able to utilize pentose substrates
naturally. Conversely, those that are able to utilize pentose sugars, such as Pichia stipitis,
produce end-products at unacceptably low yields and productivity levels (Agbogbo et al., 2006).
Strategies for enhancing the utilization of xylose and, to a lesser extent, arabinose have to date
involved such processes as genetically engineering S. cerevisiae to allow xylose utilization
(Pitkänen et al., 2005; Hahn-Hägerdal et al., 2007). Some limitations of these methods include
difficulties in using engineered microbes in large scale industrial processes, such as poor public
perception, the possibility of back-mutation and the unsuitable physiological properties of the
resulting strains. Some success has been achieved with natural selection, developing strains of
S. cerevisiae that are able to utilize xylose (Attfield and Philip, 2006; Govindaswamy and Vane,
2007). While the growth rates achieved were quite slow, the principle provides an interesting
alternative to genetic engineering.
Kawaguchi et al., (2006) have attempted amino acid synthesis using Corynebacterium
glutamicum, an industrially proven bacterium, using a xylose substrate and forcing carbon flow
to ethanol production using high cell density fermentation under bacteriostatic conditions. This
approach is unproven except in defined medium conditions and also relies on genetically
engineered strains.
Other bacterial strains capable of metabolising pentoses for the production of alternative fuels
or industrial chemicals include solventogenic Clostridium sp. for production of butanol from
pentoses (Patel et al., 2006) alkane production by Vibrio furnissii (Patel et al., 2006) and lactic
acid production by acid-tolerant, thermophilic Bacillus strains or Lactobacillus pentosus (Cruz et
al., 2007).

Chapter 1 Introduction
7
The challenge is that the fractionation process for cellullosic biomass generates only cellulose
and hemicellulose, rather than completely separating the glucose polymers from the pentose
polymers. Thus, any successful process must deal with the situation where at least two sugars
may be present, and sometimes more.
This research will investigate the use of naturally occurring bacteria, isolated specifically for the
purpose of breaking down the pentose sugars naturally present in the fraction of hemicellulose
from lignocellulosic waste. The taxonomy, ecology and metabolism of these potentially valuable
organisms will also be investigated.
1.2 Research Problem and Hypothesis
The fractions of hemicellulose from lignocellulosic waste have always contained some residual
cellulose, so the sugars available for industrial fermentation will normally contain a mixture of
pentoses plus glucose. Efficient use of this fraction as a fermentation feedstock would thus
require the use of microbes that can metabolise such pentoses in the presence of glucose,
resulting in efficient use of the sugars available.
The research hypothesis, therefore, was that pentose utilizing organisms isolated from
naturally pentose enriched environments may subsequently be used to degrade pentose
sugars such as those found in hemicellulose from agricultural waste, and produce
commercially valuable by-products.
1.3 Aims and Objectives of Research
In order to prove this hypothesis, I have defined three major aims. These aims, and the
objectives required to achieve the aims, are described in the following paragraphs.
The first aim of this research was to isolate, identify and taxonomically characterize pentose-
capable organisms from waste products of agricultural processes, specifically hemicellulose-
enriched soil from sugar cane milling.
The second aim of this research was to detect and identify major end-products from pentose
and diauxie growth of the suitable isolates obtained.
The final aim was to investigate microbial populations in pentose rich habitats, using a
metagenomic approach, specifically Denaturing Gradient Gel Electrophoresis (DGGE) and
PhyloChip. This is important, in order to relate the pentose-capable isolates back to their natural

Chapter 1 Introduction
8
environment, and to establish whether the in-situ breakdown of pentose sugars is likely to occur
naturally, or whether the process would require seeding in order to initiate the intended
metabolic outcomes.
These aims will be achieved by the completion of the following objectives:
Aim 1 - Objective 1: Collect environmental samples from pentose-rich sites, particularly those
found in sugarcane growing and processing sites, and screen these samples for their
ability to utilize pentose sugars. Identify pentose-capable isolates.
Objective 2: Test pentose (xylose, arabinose and ribose) carbon sources, to determine
the growth characteristics of pentose-capable isolates in single carbon media.
Objective 3: Test pentose-capable isolates for their growth characteristics in a dual
carbon medium, using glucose plus each of the three pentose sugars separately.
Aim 2 – Objective 1: Identify the end-products of metabolism of glucose and each of the three
pentose sugars by the test organisms.
Objective 2: Identify end-products of diauxic metabolism of glucose plus each of the
three pentose sugars by the testing of organisms.
Aim 3 – Objective 1: Analyse microbial communities in sugar mill samples of water, soil, and
bagasse leachate collected from five cane-growing areas in Queensland, Australia, using
Denaturing Gradient Gel Electrophoresis (DGGE) and subsequent DNA sequencing.
Objective 2: Analyse microbial communities present in sugar mill samples (as above)
using PhyloChip analysis methods.
Objective 3: Use the two metagenomic analyses to demonstrate the complexity and
richness of a microbial community in the most pentose-rich sample (bagasse leachate),
as compared with those present in other sugar mill samples.

Chapter 1 Introduction
9
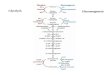
1.4 Research Plan
Aim 1: To isolate, identify and characterize pentose-capable organisms from waste products of
agricultural processes (bagasse from sugarcane)
DIAUXIE CURVES FOR GLUCOSE plus PENTOSE
GROWTH CURVES (g, µ data) for SINGLE CARBON
SOURCES
ISOLATES IDENTIFIED
SCREEN FOR PENTOSE UTILIZATION CAPACITY
COLLECT ISOLATES
Chapter 3
Chapter 4

Chapter 1 Introduction
10
Aim 2: To detect and identify major end-products from single sugar (pentose) and
diauxie(glucose plus pentose) growth of the 6 isolates obtained.
Aim 3: To investigate microbial populations in pentose rich habitats, using a metagenomic
approach, specifically DGGE and PhyloChip analysis.
END PRODUCT ANALYSIS FROM
PENTOSE METABOLISM
END PRODUCT ANALYSIS FROM MIXED SUGAR METABOLISM
IDENTIFICATION OF END PRODUCTS
COLLECT WATER, BAGASSE AND SOIL FROM SUGAR CANE
REFINERIES
ECOLOGY STUDY OF ENVIRONMENTAL PENTOSE
UTILIZING BACTERIA
(DGGE)
TAXONOMY of ENVIRONMENTAL PENTOSE UTILIZING BACTERIA
(PHYLOCHIP)
Chapter 5
Chapter 6
Chapter 7

Chapter 2 Literature Review
11
CHAPTER 2
Literature Review
2.0 Introduction
This critical review of current literature is structured as follows: firstly, a brief summary will be
presented of the history and background of industrial fermentation, leading to its particular
application for the purposes of recycling agricultural waste and production of commercially
important end-products (Section 2.1). This will be followed by a detailed analysis of the chemical
aspects of lignocellulose, in particular considering the nature of the potential substrate materials
(Section 2.2). The third section (Section 2.3) will detail the research to date, regarding the
microbial aspects of the process, including an overview of the current state of knowledge
regarding genetically altered and natural microorganisms, their end-products and ecological
aspects of research of this kind. Finally, in Section 2.4 a review of the methodology employed in
this important area of research will be reported, with particular focus on cutting-edge methods
being used in this research.
2.1 Brief Historical Background
Fermentation has always been an important part of our lives: foods can be spoiled by microbial
fermentations; and foods can be made by microbial fermentations. It was not understood how
fermentation actually worked until the research of Louis Pasteur in the latter part of the
nineteenth century and the work that followed (Laser et al., 2002). Fermentation is a microbial
process in which enzymatically controlled transformations of organic compounds occur. In this
process microbes utilize a variety of organic compounds including carbohydrates and may
produce products for use by human beings. Fermentation results in the production of foods such
as bread, wine, and beer. Bread predates the earliest agriculture and was discovered when wild
cereal grains were found to be edible (Demain, 2005). Fermented dough was thought to be
discovered by accident, when dough was not baked immediately and underwent fermentation
(Hornsey, 2003). Egypt and Mesopotamia exported bread making to Greece and Rome, and the
Romans subsequently improved the technique, leading eventually to the discovery of the role of
yeasts in baking, by Pasteur, followed by the production of baker’s yeast cultures (Hornsey,
2003).

Chapter 2 Literature Review
12
The Chinese were using fermentation around 4000 BC to produce foods such as yogurt, cheese,
fermented rice, and soy sauces. Milk has been a dietary staple since at least 9000 BC, resulting in
fermented products such as cheese, yoghurt, and sour cream (Tamang and Kailasapathy, 2010).
Modern cheese manufacturing follows almost the same steps as were used in ancient times,
including a fermentation step involving lactic acid bacteria. Many cheeses are also ripened by
means of fungi which either occur naturally or are inoculated into or onto the cheese, to add to
their distinctive flavour (Steinkraus, 1995).
Biochemically, fermentation involves the breaking down of complex organic substances into
simpler ones by means of microbial metabolic pathways. Fermentative metabolism may be
defined as the use of carbon sources to produce end-products that contain chemical energy,
usually in the absence of oxygen. These endproducts are not fully oxidized are not able to be
metabolized further in the absence of oxygen. Sugars are common substrates for fermentation
process, and examples of fermentation products are ethanol, lactic acid and lactose (Todar,
2008). In some cases the microbial cell obtains energy through glycolysis, splitting a sugar
molecule and removing electrons from the molecule. The electrons are then passed to an
organic molecule such as pyruvic acid.
Cellulosic biomass represents the only foreseeable, sustainable source of organic fuels,
chemicals, and materials (Olsson and Hahn-Hägerdal, 1996; Lynd et al., 2001). A primary
technological challenge in biologically processing cellulosic biomass into fuels and chemicals is
that of overcoming the recalcitrance of cellulose to hydrolysis. Cellulose hydrolysis processes are
typically categorized into those that use strong mineral acids and those that use cellulase
enzymes. Although processes using acids are more technologically developed, enzymatic
processes have comparable projected costs and are expected to enjoy an increasing cost
advantage as the technology improves (NREL, 1999; Lynd et al., 2001). Due to its resistance to
enzymatic attack, however, naturally occurring cellulosic biomass must be pretreated before it
can be enzymatically hydrolyzed. Pretreatment is one of the most expensive and least
technologically mature unit operations in lignocellulosic conversion processes using enzymatic
hydrolysis (Lynd et al., 2001).
To be effective, a pretreatment process must produce reactive fibre, preserve the utility of the
pentosan (hemicellulose) fraction, and limit the extent to which the pretreated material inhibits
growth of the fermenting microorganisms. To be economical, the process should minimize
energy demands and limit costs associated with feedstock size reduction, materials of
construction, and treatment of process residues Lynd et al., (2001). These process conditions

Chapter 2 Literature Review
13
such as temperature, reaction time, pH, and biomass concentration affect these substrate
factors, and thus influence pretreatment performance.
Pretreatment processes can be loosely grouped into three categories: physical, chemical, and
hydrothermal. Physical pretreatments, which are typical, demand large amounts of energy and
are expensive, employing purely mechanical means to reduce feedstock particle size so as to
increase available surface area. A variety of chemicals, such as acids, alkalis, organic solvents,
oxidizing agents, supercritical fluids, and ligninase enzymes have been considered for use as
chemical pretreatment agents (Nathan-Mosier et al., 2005). These agents are used to initiate
chemical reactions in order disrupt the biomass structure.
Hydrothermal pretreatment refers to the use of water as liquid or vapour or both, to provide the
heat to pretreat biomass. Relative to dilute acid pretreatment, hydrothermal pretreatment
processes have several potential advantages in particular the fact that there is no requirement
for purchased acid, for special noncorrosive reactor materials or for preliminary feedstock size
reduction (Nathan-Mosier et al., 2005). Furthermore, hydrothermal processes produce much
lower quantities of hydrolyzate neutralization residues, which result from the process and may
be an adverse influence on the formation of large amounts of biomass. Review of further studies
and discussion regarding microbial fermentation will be found in Section 2.3 - Microbial Aspects
of Fermentation Technology.
2.2 The Chemistry of Plant Biomass
Plant biomass is biodegradable and serves as a good alternative source of energy and
chemical products because of its safety, reliability and resulting reduction of pollution.
Rising costs, the finite nature of fossil fuels and the ecological problems associated with CO2
emissions, are combining to create renewed interest in plant biomass as a sustainable basis
for the production of alternative resources for energy, transport fuels and chemicals (U.S.
Department of Energy, 2005).
Sugar cane bagasse is the fibrous matter that remains after sugarcane is crushed to extract the
juice. A typical chemical analysis of bagasse is (on a washed and dried basis): cellulose 45–55%,
hemicellulose 20–25%, lignin 18–24%, ash 1–4%, waxes <1%. Currently bagasse is being used as
a primary fuel for sugar milling and processing operations, with occasional supplementation by
sawdust, coal and fuel oil (Womersley, 2006). The component of bagasse that is of most interest
in an industrial sense is the fraction of hemicellulose-.
The major polysaccharides in the primary wall of plants are cellulose, hemicellulose and pectin.
The secondary cell wall consists of woody tissue, which is composed predominantly of cellulose,

Chapter 2 Literature Review
14
lignin, and hemicellulose. Together, these layers are called “lignocellulose” (Almersheim, 1976 ).
Plant biomass consists mainly of three components: starch; pectin; and lignocellulose. Each of
these will now be discussed in turn, with particular respect to their potential ability to act as a
source of energy and essential chemicals.
2.2.1 Starch
Starch is the major carbohydrate reserve in plant tubers and seed endosperm where it is found
as granules, each typically containing several million amylopectin molecules accompanied by a
much larger number of smaller amylose molecules (Shigechi et al., 2004). The structure of starch
is shown in Figure 2.1. By far the largest commercial source of starch is corn (maize) with other
commonly used sources being wheat, potato, tapioca and rice. Recent years have seen the
introduction of large scale processing for the bioconversion of biomass resources, especially
starchy materials, to ethanol, which is expected to find a wide range of applications, including
use as a biofuel and as the starting material for various chemicals. However, the process is
expensive. There are two main reasons for the present high cost: firstly that the yeast
Saccharomyces cerevisiae cannot utilize raw starchy materials, so large amounts of amylolytic
enzymes, (glucoamylase and α-amylase) need to be added; and secondly that the starchy
materials need to be cooked at a high temperature (140°C to 180°C) to obtain a high ethanol
yield. Non-cooking and low-temperature cooking fermentation systems, used to reduce the
energy cost for the cooking of starchy materials, have succeeded in reducing energy
consumption of the process by approximately 50%. However, it is still necessary to add
amylolytic enzymes to hydrolyse the starchy materials to glucose (Shigechi et al., 2004) which is
a costly exercise. On the other hand, some scientists disagree with the use of starch to produce
ethanol or any other industrial chemicals from starchy crops because this practice can be seen to
be contributing to a global food shortage (Fong, 2008).
The main drawback of using starch is that its high sensitivity to moisture renders it unsuitable for
many applications. In an effort to overcome this problem, synthetic plastics have been
experimentally combined with starch (Janssen and Moscicki, 2009), leading to new materials
with properties similar to the modern plastics that fulfil the market demand. Such "mixed"
plastics may contain as much as 50% w/w starch.

Chapter 2 Literature Review
15
Figure 2.1: Chemical structure of starch.
(Source: Unina, 2009, http://www.whatischemistry.unina.it/en/starch.html)
2.2.2 Pectin
Pectin is a heteropolysaccharide compound which consists of primary cell wall of plants. Pectin
consists of a chain-like configuration of D-galacturonic acid which makes pectin-backbone
named homogalacturonan (Figure 2.2).
Figure 2.2: Chemical structure of pectin
(Source: Cybercolloids, http://www.cybercolloids.net/library/pectin/introduction-pectin-
structure)
Many plants including legumes, vegetables, and citrus fruits have high levels of pectin. Pectin is
an insoluble protoprotein which is contained in a section of non woody parts of the plants.
Pectin is considered an important dietary fibre, and is reputed to have many health benefits.
Pectin does not, however, have an important role as a nutritional or fermentable substrate. The
quantity, structure and chemical constitution of pectin in plants vary throughout the plants’

Chapter 2 Literature Review
16
structure. For example, the skins of citrus fruits, apples and other fruits contain much more
pectin than the pulp. Fruits become ripe and soften due the enzymatic action of pectinase and
pectinesterase breaking down pectin (Cybercolloids; Belitz et al., 2004).
2.2.3 Lignocelluloses
Plant cell walls are composed of cellulose and hemicellulose, pectin and in many cases lignin.
Lignocellulosic biomass refers to plant biomass that is composed of cellulose (44%),
hemicellulose (30%), and lignin (26%) (U.S. Department of Energy, 2005). The dry mass
composition of various lignocellulosic materials including carbohydrate and non-carbohydrate
are shown in Table 2.1.
Figure 2.3: Structure of lignocellulose (ref. Rubin, 2008, page 843 figure 2)
For use as a fermentation substrate, lignocellulosic biomass is firstly pre-treated by physical and
chemical means to free the polymeric components lignin, cellulose and hemicellulose.
Fractionation and enzymatic treatment can subsequently yield various output streams that are
rich in phenolics from lignin, glucose from cellulose and pentoses (mainly xylose and arabinose)
from hemicellulose. Fractionation is necessary to enable the extraction of chemicals from these
streams for further processing and the use of unfractionated lignocellulosic can lead to the

Chapter 2 Literature Review
17
inhibition of microbes during biofuel fermentations, mainly caused by phenolics and their
derivatives (Singh and Mishra, 1995).
The principal components of lignocellulose (lignin, cellulose and hemicellulose) are examined in
more detail in the following paragraphs, particularly with respect to their potential as a
substrate for microbial industrial processing.
Table 2.1: Comparison of various lignocellulosic raw materials (Lee, 1997)
Carbohydrate (% of sugar equivalent)
Corn
stover
Wheat
straw
Rice
straw
Rice
hulls
Bagasse
fibre
Cotton
gin
trash
Newsprint Populous
tristis
Douglas
fibre
Glucose 39.0 36.6 41.0 36.1 38.1 20.0 64.4 40.0 50.0
Mannose 0.3 0.8 1.8 3.0 n/a 2.1 16.6 8.0 12.0
Galactose 0.8 2.4 0.4 0.1 1.1 0.1 n/a n/a 1.3
Xylose 14.8 19.2 14.8 14.0 23.3 4.6 4.6 13.0 3.4
Arabinose 3.2 2.4 4.5 2.6 2.5 2.3 0.5 2.0 1.1
Non carbohydrate (%)
Lignin 15.1 14.5 9.9 19.4 18.4 17.6 21.0 20.0 28.3
Ash 4.3 9.6 12.4 20.1 2.8 14.8 0.4 1.0 0.2
Protein 4.0 3.0 n/a n/a 3.0 3.0 n/a n/a n/a
2.2.3.1 Lignin
Lignin is a complex polymer of phenylpropane units and is a component of wood. It is cross-
linked to cellulose fibers, hardening and strengthening the cell walls of plants with a variety of
different chemical bonds. The function of lignin is to give plants structural rigidity and to protect
their cellulose and hemicellulose from microbial attack. It has been shown that the fungi and
actinomycetes are able to degrade very low amount of lignin (Richard, 2006). This is relevant for
microbial fermentation processes. However, since lignin is very difficult to degrade, it is unlikely
to be a good choice for use in industrial fermentation processes.
2.2.3.2 Cellulose
Cellulose is a polysaccharide which contain a straight chain of few hundred to over ten thousand
β (1→4) linked D-glucose molecules. There are only few enzymes are needed to break down this

Chapter 2 Literature Review
18
molecule due to its conservative structure and repeated similar linkages. Humans do not have
suitable enzymes to break down cellulose but some microorganisms are capable of degrading
cellulose.
Traditionally, baker’s yeast (Saccharomyces cerevisiae), has been used in the brewery industry to
produce ethanol from hexoses (6-carbon sugars). Due to the complex nature of the
carbohydrates present in lignocellulosic biomass, a significant amount of xylose and arabinose
(5-carbon sugars derived from the hemicellulose portion of the lignocellulose) is also present in
the hydrolysate. For example, in the hydrolysate of corn stover, approximately 30% of the total
fermentable sugars is xylose (Karhumaa et al., 2005). As a result, the ability of the fermenting
microorganisms to use the whole range of sugars available from the hydrolysate is vital to
increase the economic competitiveness of cellulosic ethanol and potentially bio-based
chemicals.
Recently, genetically engineered yeasts has been developed to efficiently ferment xylose and
arabinose (Öhgren et al., 2006), and even both together. Yeasts are especially attractive for use
in cellulosic ethanol processes as they have been used in such processes for hundreds, even
thousands, of years. Their advantages are that they are tolerant to high ethanol and inhibitor
concentrations and as they can grow at low pH values. This prevents bacterial contamination
(Karhumaa et al., 2005), bacteria being less tolerant of acidity than yeasts.
2.2.3.3 Hemicellulose
Hemicellulose are a heterogeneous class of polymers representing in general, 15–35% of plant
biomass and which may contain many different sugar monomers: pentoses (d-xylose, l-
arabinose); hexoses (d-mannose, d-glucose, d-galactose); and/or uronic acids (d-glucuronic, d-4-
O-methylgalacturonic and d-galacturonic acids). Hemicellulose contain mostly d-pentose sugars
and occasionally small amounts of L-sugars as well. Xylose is always present in the largest
amounts, but mannuronic acid and galacturonic acid also tend to be present. Other sugars such
as l-rhamnose and l-fucose may also be present in small amounts and the hydroxyl groups of
sugars may be partially substituted with acetyl groups (Desantis et al., 2007). Xylans are the
main hemicellulose components of secondary cell walls constituting about 20-30% of the
biomass of hardwoods and herbaceous plants. Mannan-type hemicellulose like glucomannans
and galactoglucomannans are the other two major hemicellulosic components of the secondary
wall of softwoods whereas in hardwoods they occur in minor amounts. In some tissues of
grasses and cereals, xylans can account for up to 50% (Heinze et al., 2005). Xylans are usually

Chapter 2 Literature Review
19
available in huge amounts as by-products of forest, agriculture, agro-industries, wood pulp and
paper industries.
In contrast to cellulose, which contains only anhydrous glucose, sugar monomers in
hemicellulose can include not only glucose, but also xylose, mannose, galactose, rhamnose, and
arabinose. Unlike cellulose, hemicellulose (also a polysaccharide) consists of shorter chains -
500-3000 sugar units as opposed to 700- 15,000 glucose molecules per polymer present in
cellulose. In addition, hemicellulose is a branched polymer, while cellulose is unbranched. Xylan
is an example of a pentosan consisting of D-xylose (Figure 2.4) units with 1β→4 linkages.
Figure 2.4: Chemical structure of xylan and xylose formation (Held, 2012, page
3, Figure 3)
Utilization of the fraction of hemicellulose in lignocellulosic biomass is an important factor in
optimizing the economics of biomass fuel processes, given that hemicellulose makes up a
significant proportion of the potentially available carbon for fermentation or chemical
extraction. Although there are several potential uses of hemicellulose-enriched fractions as
specialty or industrial chemicals produced by non-biological processes, the use of hemicellulose
carbon as a fermentable substrate remains an attractive route for producing chemicals that have
an intrinsic commercial value.
2.3 Microbial Aspects of Fermentation Technology
2.3.1 Fermentation of lignocellulosic sugars
Research and development studies of fermentation technology have been conducted over the
last few decades to make fermentation processes more efficient. In principle, most of the

Chapter 2 Literature Review
20
biochemical and microbiological problems, such as discovering suitable strains of
microorganisms, and establishing optimal conditions, have been addressed.
Fermentation of lignocellulosic biomass is a good option to deal with several industrial and
agricultural by-products. Lignocellulosic biomass is a sustainable product. There are several
energy crops that might be cultivated at different seasons of the year, and these could deliver
high energy industrial products throughout the year, rather than seasonally (Lin and Tanaka,
2006).
However, the problem still to be solved is that of identifying suitable pentose utilizing
microorganisms. Unlike the age-old processes based on Saccharomyces fermentation of glucose,
processes and organisms for use with lignocellulosic feed-stock are yet to be put into practice in
an industrial sense, and are still in the research and development phase. Some of the difficulties
encountered in this area of research will be now discussed.
The first issue to be resolved in studying the effect of pentose fermentation pathways is an
observed inhibition effect. Castro et al., (2003) reported that in order to minimize negative
effects on the pentose fermentation process, it was necessary to: prevent accumulation of
inhibitory components (such as hydroxymethylfurfual) by detoxifying them; improve media
composition; and allow the microorganism to acclimatize to the toxic inhibitors. In addition,
hemicellulosic hydrolysates are used to dilute the raw material to a standard sugar
concentration. Similar processes may be applied for the industrial production of ethanol. This
has the potential to ensure high levels of product whilst avoiding treatment designed to remove
inhibitory compounds that are normally found in the hemicellulosic acid hydrolysate. Such a
dilution step also decreases the necessity for addition of extra microbial nutrients and requires
minimal changes to industrial fermentation plants and processes for its implementation. The
performance of an innovative two-stage continuous bioreactor with a cell recycle-potential,
capable of giving very high productivity was investigated by Chaabane et al., (2006).
Fermentative activity of the yeast, Saccharomyces was not influenced by the total biomass
concentration in the range tested (up to 157 g L−1). A key parameter for improving ethanol
production would then be a better management of the cell viability (Chaabane et al., 2006).
A second issue related to the effective utilization of lignocellulosic biomass by fermentation is
the presence of a mixture of carbon sources, the major components being cellulose, which is
almost all glucose, and hemicellulose, which consists partly of pentose and partly of hexose
sugars. Consequently, pentose sugars (e.g., xylose, arabinose, and ribose) constitute a smaller
proportion of lignocellulosic biomass than do hexoses. Moreover, microorganisms generally use

Chapter 2 Literature Review
21
hexoses preferentially over pentoses. Therefore, reliance on the fermentation of pentoses alone
could never become a viable strategy for commercial fermentation processes. Inevitably, there
are some exceptions, such as the conversion of xylose to the sweetener xylitol, but such cases
are rare (Hahn-Hägerdal et al., 2007).
Unfractionated hydrolysates, including pentoses, would obviously be the preferred fermentation
substrate so long as the microorganism can convert the various available sugars, whether they
are pentoses or hexoses or a mixture of these (Bruinenberg et al., 1983). Culture-based solutions
to the mixed carbon-sources problem include sequential fermentation with different microbial
species, and the co-culture of several microorganisms with different substrate capabilities.
Alternatively, the use of a single microorganism capable of using all of the substrates present is
an attractive option. Due to the resulting simplification of the industrial process a single-vessel
fermentation step may be employed, with a single starter culture as the inoculum.
One very common outcome of mixed carbon source fermentation is the diauxie phenomenon,
whereby one sugar (generally the energy-efficient glucose) is used prior to the other(s). This
characteristic can be easily observed by measuring the growth curves of an organism growing in
a mixed carbon source medium, noting particularly that two distinct growth phases are present,
separated by a lag period. The diauxie growth pattern is generally attributed to catabolic
repression (Nishizawa et al., 2004).
Inhibition of β-galactosidase expression in a glucose-lactose diauxie system is a typical example
of a catabolic repression effect in Escherichia coli, and this has therefore been used as a model
system to investigate the mechanism of the diauxie phenomenon. It is assumed that the
inhibitory effect of glucose on the expression of the lac operon is mediated by a reduction of the
cyclic Adenosine Mono Phosphate (cAMP) levels in the glucose-lactose system (Grimmler et al.,
2010). However, there is no direct evidence to support this explanation.
In order to examine the roles of cAMP and the cAMP receptor protein (CRP) in the diauxie effect,
the intracellular levels of these factors were determined during diauxie growth in a glucose-
lactose medium (Verho et al., 2003; Wang et al., 2005). The levels of cAMP and CRP in the
lactose grown phase were not higher than those in a glucose-grown phase. Addition of
exogenous cAMP eliminated diauxie growth but did not eliminate glucose repression. Glucose
repression and diauxie were observed in cells that lacked cAMP but produced a cAMP-
independent CRP (Wong et al., 1997; Jeppsson et al., 2006). In addition, inactivation of the lac
repressor by the disruption of the lacI gene or the addition of IPTG, effectively eliminated

Chapter 2 Literature Review
22
glucose repression. For this purpose, a novel lag model was proposed for diauxie, which has two
functional forms, each embodying the dependence of total cell mass on the lag period with
higher cell mass resulting in a shorter period of growth suppression between primary and
secondary substrates (Wong et al., 1997).
The efficiency of sugar breakdown in microorganisms is dependent not only on the metabolic
pathways available to the organism, but also on the effective transport of the sugar molecules
into the microbial cell. Carrier-mediated transport is the predominant mechanism of sugar
uptake in microorganisms. Specific mechanisms and relative uptake rates for various sugars are
species dependent (Singh and Mishra, 1995). At least in baker’s yeast (S. cerevisiae), the uptake
rates of xylose and glucose are comparable, but the efficiency of xylose transport appears to be
lower than that of glucose, and the yeast’s affinity (Michaelis constant) for xylose is much less
than for glucose (Singh and Mishra, 1995; Toivari et al., 2001).
Efficient use of the lignocellulosic fraction as a fermentation feedstock may be effected by one
of at least three possible scenarios. The first option requires microbes that can metabolise the
pentoses in the presence of glucose, preferably without being subject to catabolite repression
and so being capable of the simultaneous use of the sugars. A second possibility is that the
selected microorganism should carry out an efficient diauxie process using two or more sugars
sequentially. A third option is to use a sequential process involving a number of microbial
processes to enhance the selective fermentation of hemicellulose pentoses in a mixture of
substrates. In such a scenario, any glucose present could be used later, or earlier, in ethanol
production by S. cerevisiae (Attfield and Philip, 2006). This is an attractive option, because large
scale ethanol production by S. cerevisiae is an established technology that will be hard to replace
by alternative biological processes (Becker and Boles, 2003). From the perspective of maximizing
the yield of ethanol as a biofuel, a process that can rescue glucose from the hemicellulose
fraction for use in high yielding yeast fermentation would be desirable (Hahn-Hägerdal et al.,
2006), and would consequently prevent the possibility of catabolite repression.
2.3.2 Microorganisms with a natural ability to ferment lignocellulosic biomass
Several microorganisms, including certain bacteria, yeasts and filamentous fungi, have been
reported as being able to ferment lignocellulosic hydrolysates, so generating ethanol. As ethanol
production is the oldest and most familiar industrial fermentation process known, this is the best
researched of the microbial fermentation processes.
It might reasonably be expected that a pentose-utilizing microorganism can be found in a
hemicellulose-enriched habitat. Such environments include any areas where decaying plant

Chapter 2 Literature Review
23
matter may be present, such as soil and natural water-sources (ponds, streams). Soil bacteria
such as Mycobacterium spp, Corynebacterium sp, and Clostridium acetobutylicum, are able to
utilize pentoses naturally (Singh and Mishra, 1995). Others mentioned in the literature have
included bacteria such as Klebsiella species and Bacillus sp, and yeasts such as Pichia stipitis, and
Kluveromyces species. A number of other naturally occurring organisms that convert pentoses to
ethanol are listed in Table 2.2 (Singh and Mishra 1995). Kastner et al., (1999) listed the yeast P.
stipitis, Pachysolen tanophilus and Candida shehatae as potential pentose fermenters.
Rhodococcus spp are particularly important due to their ability to catabolize a wide range of
compounds and produce bioactive steroids, acrylamide and acrylic acid and their involvement in
fossil fuel biodesulfurization (McLeod et al., 2006).
Table 2.2: List of pentose utilizing microorganisms (adapted from Singh and Mishra, 1995)
The common and well-known bacterium, Escherichia coli, also has some advantageous
characteristics as a candidate for ethanol production, such as its ability to ferment a wide range
of sugars, including D-xylose and L-arabinose, its capacity for simple genetic manipulation and its
history of prior industrial use (e.g. for the production of recombinant proteins). However, a
number of drawbacks can also be identified: the neutral preferred pH range (6–8) makes
bacterial fermentation susceptible to contamination; the low tolerance to lignocellulose derived
inhibitors; low ethanol tolerance; and mixed product formation (ethanol, acetic acid, lactic acid
and others), reducing the yield of any single product such as ethanol (Dien et al., 2003).
The high ethanol yield and specific productivity observed for the yeast, Zymomonas mobilis are
consequences of anaerobic D-glucose utilization using the Entner–Doudoroff (ED) pathway.
However, Z. mobilis has disadvantages similar to those of E. coli, such as the neutral pH range
and low tolerance to lignocellulose-derived inhibitors. Moreover, Z. mobilis has a narrow
substrate range, lacking the ability to utilize all of the main sugars from lignocellulose except D-
glucose.
Bacteria Yeasts and moulds Actinomycetes
Zymomonas mobilis Zymobacter palmae Escherichia coli Salmonella typhimurium Clostridium acetobutylicum Bacilus subtilis
Saccharomyces cerevisiae Rhodotorula glutinis Pichia stipitis Candida shehatae Aspergillus niger Rhizopus solani Fusarium oxysporum
Rhodococcus

Chapter 2 Literature Review
24
The yeast S. cerevisiae is the most commonly used microorganism in traditional industrial
fermentations, including current sucrose, starch and cellulose based fermentation processes,
and can efficiently ferment simple hexose sugars, such as D-glucose, D-mannose and D-
galactose, and disaccharides like sucrose and maltose (Richard et al., 2003). S. cerevisiae also has
a relatively good tolerance to lignocellulose-derived inhibitors and high osmotic pressure
(Moorbach and Krämer, 2003). Furthermore, the fermentation rate is not significantly reduced
at ethanol concentrations below 10% (v/v) (Ozcan et al., 1991). The major inconvenience in
using S. cerevisiae for lignocellulosic fermentation is its lack of any natural ability to utilize the
pentose sugars D-xylose and L-arabinose, which dominate the pentose sugar fraction of
hemicellulose.
In contrast to S. cerevisiae, the yeast Pichia stipitis is able to metabolise the main hemicellulose
sugar monomers and to ferment xylose. P. stipitis, its anamorph Candida shehatae, and also
Pachysolen tanophilus efficiently ferment xylose but only under oxygen limited conditions
(Ligthelm et al., 1988). Moreover, these yeasts are less tolerant to pH, ethanol and hydrolysate
inhibitors when compared to S. cerevisiae (Hahn-Hägerdal et al., 2007). P. stipitis is also
described as being capable of producing hemicellulolytic enzymes (Jeffries et al., 2007).
2.3.3 Genetic modification of microorganisms to allow pentose fermentation
A lack of microorganisms that are able to naturally and efficiently ferment hexoses and pentoses
is a major constraint to the economic utilisation of biomass. Therefore, recombinant strains of
bacteria and yeast have been developed to meet the requirements of industrial lignocellulose
fermentation.
Escherichia coli, Klebsiella oxytoca, and Zymomonus mobilis have all been genetically engineered
to produce ethanol efficiently from all hexose and pentose sugars present in the polymers of
hemicellulose (Ingram et al., 1987; Dien et al., 2002; Kim et al., 2007; Yanase et al., 2007). Three
recombinant fermentation strains were designed as candidates for the improvement of biofuel
processes. These were S. cerevisiae, E. coli and Zymomonas mobilis ( Singh and Mishra, 1995;
Hahn-Hagerdal et al., 2001; Attfield and Philip, 2006). Others mentioned in the literature
included Klebsiella species, Pichia stipitis, Bacillus species and Kluveromyces species (Bothast et
al., 1999).
Kawaguchi et al., (2006, page 3418) reported that wild type “C. glutamicum was unable to utilize
xylose under both standard aerobic and oxygen deprivation conditions, owing to the lack of
xylose isomerase activity. However, metabolically engineered C. glutamicum has been widely

Chapter 2 Literature Review
25
used for the industrial production of various amino acids and nucleic acids from sucrose and
glucose based media”.
In my opinion, both biocatalyst and genetically modified derivatives of microbes are potentially
useful for the conversion of pentose-rich feed stocks such as corn stover, corn fiber, bagasse,
rice hulls, and rice straw into commodity chemicals such as lactic acid, fatty acids, acetic acid and
several other industrial chemicals. However the genetic modification processes are time
consuming and expensive.
2.3.4 End-products of microbial fermentation of lignocellulose
According to Rosenberg (1980), methods are available for biologically converting pentose sugars,
which are the major constituents of hemicellulose, to ethanol and other neutral products. These
products could be used as clean-burning liquid fuels, solvents or chemical feed-stocks. Pentose
fermentations which did not yield ethanol as a significant product (>5% yields, g/g substrate
fermented) were not considered as potential biofuel processes (Rosenberg, 1980). Lactic acid,
fatty acids and acetic acid are used in the food industry as additives, flavouring agents and
preservatives. A list of microbial end-product from pentose fermentation is supplied in Table 2.3.
The production of lactic acid and its derivatives is economically dependent on many factors of
which the cost of raw material is highly significant. It is very expensive to use purified sugars like
glucose and sucrose as the feedstock for lactic acid production. Different food or agro-industrial
products or residues are cheaper alternatives to refined sugars for lactic acid production.
Sucrose-containing materials such as molasses, starchy materials and lignocellulose/
hemicellulose hydrolyzates are most economical for use in lactic acid production (Patel et al.,
2004). Sugarcane bagasse has been reported to be useful in lactic acid production by the fungus
Rhizopus oryzae and the bacterium Lactobacillus. However, it was used only as a supplement to
the sugars or starch hydrolyzate that comprised the major carbon source (Barker and Worgan,
1981). Generally, Lactobacillus species are deficient in cellulolytic and amylolytic capacity (that
is, they lack the capability for breaking down starch into sugars), so necessitating the prior
hydrolysis of cellulosic and starchy wastes to improve their utilization (John et al., 2007).
Corynebacterium spp are Gram-positive bacteria, constituting part of the Actinomycetes
subdivision of Eubacteria. Some unknown pathogens of human and others are industrially
important due to their ability to degrade a wide range of chemicals. When grown in a mineral
medium and under conditions of oxygen deprivation, this aerobic bacterium is essentially under
bacteriostatic conditions, but continues to maintain its major metabolic capabilities. It is
therefore able to excrete significant amounts of several metabolites, such as lactic, succinic, or

Chapter 2 Literature Review
26
acetic acids, even though cellular growth is essentially arrested (Kawaguchi et al., 2006). By
stopping the production of cell biomass a process will occur, which stimulates the productiony of
end products. . C. glutamicum uses many ordinary substrates such as sucrose and glucose for
industrial product yield. But until recently C. glutamicum were unable to utilize xylose due to
lack of appropriate enzyme systems. Xylose degradation is necessary for microorganisms that
are required to produce valuable industrial products from lignocellulosic biomass. Kawaguchi et
al., (2006) solved this problem by modifying a recombinant C. glutamicum strain with ability to
utilizing both glucose and xylose.Using the conventional, well developed methods of thermal or
acidic pre-treatment (Lee, 1997), lignocellulosic raw materials are de-lignified. Then, either
simultaneously (simultaneous saccharification and fermentation process) or separately, ethanol
fermentation is carried out utilizing the released hexose and pentose sugars in the cellulose and
hemicellulose hydrolyzates.
Scientists at the Solar Energy Research Institute, USA (U.S. Department of Energy, 2005)
conducted an economic analysis of xylose fermentation and a simultaneous saccharification and
fermentation process for wood to ethanol conversion. It was demonstrated by this team, that
the production cost of ethanol could be reduced from $US1.65 to $US1.23 per gallon (converted
to Australian currency (exchange rates correct as on 11th July 2012).This is a reduction of 1.61
cents/litre to 1.20 cents/litre) if all the xylose was utilized for the ethanol production. Enhancing
the yield of xylose utilization from 85% to 95% resulted in a cost reduction by 2.4 cents (US) per
gallon, and in a decrease in fermentation time from 2 days to 1 day, bringing about a cost
reduction of 1.3 cents (US) per gallon. Based on the cost of wood at $US34 per dry ton and on a
fermentation capacity of 10,000 tons per day, an estimated ethanol production cost of 74 cents
(US) per gallon (21 cents (AU$) per L) was obtained. From this information, it is evident that
bioconversion of xylose to ethanol has the potential to markedly affect the cost of overall
ethanol production from lignocellulosic biomass (Von Sivers and Zacchi, 1996).

Chapter 2 Literature Review
27
Table 2.3: List of microbial products from pentose fermentation (Singh and Mishra, 1995)
Product name Bacteria Yeast
Acetic acid, Acetone,
butanol, Propionic acid
Clostridium, Bacillus
Lactic acid, Citric acid Bacillus Aspergillus, Saccharomyces,
Candida
Xylitol, Pentitol Corynebacteria,
Mycobacterium
Saccharomyces, Pichia, Candida
In terms of non-ethanol end-products, amino acids are amongst the most important, particularly
in nutrition, owing to their central role in biochemistry. Convenient sources of amino acids are
being researched for industrial production purposes. For example, Beaman et al., (1971 ) were
able to recover more than trace amounts of arginine, aspartic acid, glycine, lysine, serine,
threonine, valine, phenylalanine, leucine, and isoleucine from the cell walls of Nocardia rubra.
They also confirmed the presence of minor amounts of several amino acids in thoroughly
washed and unextracted cell walls of Nocardia spp. It was observed that alkaline ethanol
removed these "background amino acids" too.
Some investigators have suggested that the amino acids found in small quantities in cell wall
hydrolysates are the result of contamination with cytoplasmic components. It would appear
from a quantitative investigation, using both alkaline ethanol extracted and unextracted cell
walls prepared from actively growing bacteria (N. Rubra) (Beaman, 1975), that a significant
amount of peptide is associated with the lipoidal components of the outer cell envelope. The
loss of this material is associated with a loss of a characteristic pattern of convolutions present
on the surface of actively growing N. rubra. Earlier investigators, employing growth conditions
differing variously from those employed by Beaman et al., (1971) and using mild methods of
extracting carefully prepared cell walls, noted the effects of such extraction on the
ultrastructures of the cell envelope.
According to Hermann (2003), the total annual worldwide consumption of amino acids in 2003
was estimated to be over 2 million tons (1,814,370 tonnes), a figure which has no doubt been
exceeded in 2012. The annual demand for amino acids like MSG-based flavour enhancers or feed
additives comprised mainly of L-lysine, D, L-methionine and L-threonine, is estimated to be
significantly higher than 1 million tons (907,185 tonnes) each. In addition, the annual demand

Chapter 2 Literature Review
28
for amino acids used in pharmaceutical products, mainly for intravenous nutrition, was 15,000
tons (13,608 tonnes).
l-Valine,l-isoleucine, l-threonine, l-aspartic acid and L-alanine are some of the amino acids
produced by Corynebacterium. Around 1.5 million tons (1.36 million tonnes) L-glutamic acid are
produced per year using coryneform bacteria. It is clear that amino acids are essential
requirements for the food and pharmaceutical industries and that the Corynebacterium group of
bacteria are capable of producing amino acids. Since amino acids are essential requirements for
the food and pharmaceutical industries, it follows that the coryneform group of bacteria should
prove to be an ideal group for the purpose of synthesising those products from previously
discarded agricultural waste, which is the main focus of my research.
2.3.5 Ecology of microbial communities
Microbes are capable of changing their habitat, metabolic characteristics, and ecological roles
over time. It is possible to detect microbial roles and densities in the environment by ecological
analysis. Fast and reliable cultivation-independent identification techniques for filamentous
bacteria in activated sludge have been employed for the evaluation of their importance in the
sewage treatment process as well as for investigation into the problems that may arise during
the process. The earlier study by Schuppler et al., (1998) demonstrated that the classical
approaches failed to differentiate foam-causing species such as Gordona amarae, Rhodococcus
rhodochrous and Tsukamurella paurometabolum due to their variable morphology, staining
behaviour and fastidious nature. As previous research has shown, there are several methods
available for comparative sequence analysis of 16S rRNA sequences directly retrieved from
natural microbial communities. This represents the most powerful method for describing species
composition of ecological niches, as both cultured and as yet uncultured microorganisms can be
identified.
Microbial ecology research is generally based on either autecological or synecological
approaches. Autecological studies are those involved with the behaviour of the individual
species with in a population, while synecological studies deal with interaction of populations of
different kinds of microorganisms within an ecosystem (Craig et al., 2010). Survival and activity
of viruses and bacteria in sewage systems, surface water, groundwater, and aerosols are
examples of suitable issues for synecological study. Indicator organisms, such as E. coli, in
sewage water and receiving water fall in the autoecological domain (NH&MRC, 2004).
Autecological study of indicator bacteria involves the issues of the habitats, life cycles and

Chapter 2 Literature Review
29
nature of the relationship of the indicator to the presence of pathogenic bacteria (Craig et al.,
2010).
Habitatoriented issues are generally best studied by synecological methods, Example include
studies of biogeochemical cycling (the carbon cycle, nitrogen cycle, sulphur cycle etc.), microbial
ecology of the rumen, microbial interactions in sewage, biodegradation of pollutants in soils and
aquatic ecosystems, and the production of goods such as cheese, pickles and fermented
beverages (Christon, 1991).
Metagenomic libraries with molecular bases (phylogenetics, DNA microarrays, and functional
genomics) are powerful tools for exploring microbial diversity in various ecosystems, and also for
investigating the large amount of data regarding the genetic information derived from
uncultured microorganisms. This information will form the basis of new initiatives aiming to
conduct genomic studies that link phylogenetic and functional information about environmental
microbial flora, including both culture-capable and uncultured.
Craig et al., (2010) reviewed progress toward understanding the biology of uncultured Bacteria,
Archaea, and viruses through metagenomic analyses. For sequence-based approaches, the
speed and cost of nucleotide sequencing will cease to be a barrier as sequencing technology
continues to improve and becomes less expensive. Advances that will facilitate the management
and analysis of large libraries include bioinformatics tools to analyze vast sequence databases
and reassemble multiple genomes rapidly, together with affordable gene chips for library
profiling and the ready distinction of clones that are expressing genes, from those clones that are
silent. Functional analysis will require more innovation in method development. Most important
among these are the strategies to improve heterologous gene expression and approaches for the
efficient screening of large libraries (Sebat et al., 2003). Amann et al., (1995) estimated that
>99% of microorganisms that were observable in nature typically were not able to be cultivated
using standard culture techniques. The majority of the bacterial taxonomic divisions are poorly
represented by cultured organisms. The overall performance of 16S rDNA sequence analysis was
considered to be excellent, since it was able to resolve almost 90% of identifications, when
applied to a large collection of phenotypically unidentifiable bacterial isolates. In order to
improve this performance, efforts should be made to complete 16S rDNA databases with high-
quality sequences and to develop electronic tools for sequence comparison and interpretation.
2.4 Methodology
2.4.1 End product analysis by high performance liquid chromatography (HPLC)

Chapter 2 Literature Review
30
The identification and analysis of amino acids has permitted vast research efforts into protein
and food testing since Moore and Stein (1951) invented an ion exchange chromatography
process to isolate un-derivatized amino acids (AAs) subsequent to post-column derivatization
with ninhydrin and further identification. Many developments followed this initial work,
resulting in the sophisticated procedures such as HPLC, which are used for the same purposes,
but with much greater efficiency and accuracy today. OPA is used to detect primary amino acids.
However, the secondary AAs were not detected, because another derivatizing agent, 9-
fluorenylmethyl chloroformate (FMOC), necessary for their detection, was not used in this
research.
High performance liquid chromatography (HPLC) is basically a highly improved form of column
chromatography. Instead of a solvent being allowed to drip through a column under gravity, it is
forced through under high pressures of up to 400 atmospheres (Lefebvre et al., 2002). There are
some differences between HPLC and gas chromatography (GC), which make HPLC cheaper and
faster than GC. Firstly, the process of separating the compounds in a mixture is carried out
between a liquid stationary phase and a gaseous mobile phase in a GC, whereas in HPLC the
stationary phase is a solid and the moving phase is a liquid. Secondly, the column through which
the gas phase in GC passes is located in an oven where the temperature of the gas can be
controlled, whereas HPLC typically has no issues with temperature control. Thirdly, for a GC the
concentration of a compound in the gas phase is solely a function of the vapour pressure of the
gas; whereas HPLC is not affected by such a concentration problem.
Liquid chromatography with electrochemical or fluorescence detection has been used to analyse
amino acids. However, the fluorescence detection has greater sensitivity and so is usually
selected. As amino acids do not fluoresce, a derivatizationis required. The reagent o-
phthalaldehyde with 2-mercaptoethanol (OPA/2-ME) as catalyst is as derivatizant (Moore and
Stein, 1951; Peris-Vicente et al., 2005). The drawback of derivatization using in amino acid
analysis can be impaired by the presence of some pigments because the metallic cations of the
pigments can form complexes with some amino acids, changing the relative amount of free
amino acid. The use of a cation sequestering reagent such as ethylendiamine tetraacetic acid
(EDTA) can avoid the interference of the pigments (De La Cruz-Canizares et al., 2004).
In experiments performed by Peris-Vicente et al., (2005) asparagine and glutamine were not
analyzed, because hydrolysis converts them to aspartic and glutamic acid. The glutamic acid is
also partially converted into pyroglutamic acid during hydrolysis. The hydrolysis destroys
tryptophane, and the o-phthalaldehyde derivatives were weak, so proline and the cysteine did
not appear.

Chapter 2 Literature Review
31
The direct UV or fluorescence detection of amino acids by chromatographic analysis is difficult
due to the absence of a strong chromophore or fluorophore. Specific detection approaches
have been applied such as amperometric/electrochemical, evaporative light scattering,
chemiluminescent nitrogen and mass spectrometry detectors (Zoppa et al., 2006). The
combination of chromatographic separation and pre- or post column derivatization followed by
UV or fluorescence detection remains the most convenient and widely used analytical approach
to improve sensitivity. In particular, pre-column derivatization offers the advantage of increasing
hydrophobicity of the amino acids so that they can be retained on the columns. Commercially
available derivatization UV and fluorogenic reagents commonly used for the pre-column
derivatization of amino acids are phenyl isothiocyanate (PITC), 4- nitrobenzoyl chloride, p-
nitrobenzyl bromide, o-phthalaldehyde (OPA), 9-fluorenylmethyl chloroformate (FMOC-Cl),
dansyl chloride (Dns-Cl), 4-fluoro-7-nitro-2,1,3-benzoxadiazole (NBD-F), and naphthalene-2,3-
dicarboxaldehyde (Peace and Gilani, 2005). However, the use of these reagents can involve
different drawbacks such as limited selectivity and sensitivity, low stability of the derivatives,
time-consuming derivatization procedure or the need for extraction procedures before the
analysis. OPA has the advantage of reacting rapidly under mild conditions with the primary
amino group, but generally it is used as a post-column derivatization reagent owing to the
instability of its derivatives.
Recently, 2,7-dimethyl-3,8-dinitrodipyrazolo 1,5-a:1′,5′-d]pyrazine-4,9-dione (DDPP) and 4,7-
phenan throline-5,6-dione (phanquinone) were proposed as new pre-column derivatization
reagents for HPLC analysis of amino acids using UV or fluorescence detection (Gatti and Gioia,
2008). Both compounds have been proven to be selective towards the amino function giving
stable derivatives, useful for quality control of commercial formulations. The high sensitivity of
the phanquinone method also allowed its application to biological samples. Gatti et al., (2010)
focused on the use of 2,5-dimethyl-1H-pyrrole-3,4 dicarbaldehyde (DPD). Owing to its structural
analogy with OPA, it has the intrinsic potential to react quickly under mild conditions with the
functional site of amino acids (Figure 2.5), but has not previously been studied as an analytical
reagent (Gatti et al., 2010).

Chapter 2 Literature Review
32
Figure 2.5: o-phthalaldehyde (OPA) and mercaptoethanol reaction (Ref: Woodward and
Henderson, 2010, page 2, figure 1)
The Agilent 1100 HPLC is used for immediate identification of amino acids by ZORBAX Eclipse
AAA high efficiency HPLC column. It has a short analysis time from injection to injection can be
as low as 14 min (10 min analysis times) for a 7.5 cm column and 24 min (16 min. analysis times)
on the 15 cm long column. Sensitivity (5 - 50 pmol with DAD, FLD) and accuracy are gained by
using both OPA and FMOC derivitization in Agilent 1100 HPLC instrument. The Eclipse AAA is
using OPA and FMOC derivatization process for quick powerful resolution of 24 amino acids
separation (Henderson, 2009; Woodward and Henderson, 2010).
2.4.2 Denaturing gradient gel electrophoresis (DGGE)
Microbial ecological analysis has been dramatically improved in the last ten years. Conventional
procedures like culture and microscopic methods are enough to superficially explore the
microbial communities in our environment. Microscopic identification is useful for many natural
bacteria due to their irregular and small morphological cell size. However, media used for the
cultivation of environmental microorganisms do not necessarily support growth of an
increasingly recognized portion of the indigenous population and so may give a biased view of
the community composition. Isolation (culture) of the vast majority of naturally occurring
bacteria in pure culture is impossible (Muhling et al., 2008), due to our lack of knowledge of the
specific culture conditions they require and due to the potential for synergy between different
organisms. Comparisons of culturable and total microscopic cell counts from diverse habitats
have demonstrated the inadequacy of the culture-dependent approach to analyse microbial
community composition effectively (Muhling et al., 2008). Therefore, other tools are required to
supplement the conventional microbiological techniques used in microbial ecology, including
those that use the gene sequences of DNA as a molecular marker for identification of

Chapter 2 Literature Review
33
microorganisms. This has dramatically changed our perception of the diversity of microbial
communities.
One of these techniques is denaturing gradient gel electrophoresis (DGGE) in which DNA
fragments are separated in a gradient of DNA denaturants according to differences in their
sequences. Temperature gradient gel electrophoresis (TGGE) is a similar process but with a
changing temperature gradient (TGGE) to distinguish the DNA fragments. Both are relatively
easy to perform and are especially well suited to the analysis of multiple samples (Ercolini,
2004). Since the introduction of DGGE into microbial ecology by Muyzer and Smalla (1998), and
later work by Brons and van Elsas (2008), this method has been adapted in many laboratories as
a convenient tool for the assessment of microbial diversity in natural samples.
DGGE methods are applied to identify the similar length of double-stranded DNA fragments
through PCR amplification. The difference between the strength of GC (3 hydrogen bonds per
pairing) and AT base pairing (2 hydrogen bonds) are detected by this method. Generally high
concentration of GC containing double-stranded DNA will be stay long till they migrate to higher
concentration of denaturation. It is preferable to travel large size of double-stranded DNA in the
acrylamide gel by electrophoresis. In this situation, differing sequence of DNA fragments could
be identified in that gel (Muyzer and Smalla, 1998).
DNA extractions from microbial communities are amplified with specific set of primers for 16S
rDNA called PCR products. This is difficult to separate from each other of same size PCR products
by agarose gel electrophoresis (Dees and Ghiorse, 2001). Sequence variations between different
bacterial rDNAs bring about different melting properties of these DNA molecules, and separation
can be achieved using polyacrylamide gels containing a gradient of DNA denaturants, such as a
mixture of urea and formamide.
The PCR products are double stranded before the gel run starts. Then the products enter into
the denaturing condition and it slowly becomes stronger as the product traverses the gel. PCR
products with different sequences therefore start melting at different positions (i.e at different
denaturant concentrations) in the gel. Melting proceeds in ‘melting domains’. Once a domain
with the lowest melting temperature reaches its melting temperature at a particular position in
the denaturant gradient, a transition from a double-stranded to a partially melted molecule
occurs. The protruding single strands effectively cause a halt in the progress of the molecule at
that position. To avoid the entire separation of two DNA strands, a 40-nucleotide GC-rich
sequence (‘GC-clamp’) is adhere at the 5’-end of each PCR primers (Horst-Backhaus et al., 1996;
Morimoto et al., 2005).

Chapter 2 Literature Review
34
PCR-DGGE is an identifying technique for observing to differentiate of microbial genetic diversity
and measuring the richness of dominant microbial species. Moreover, DGGE permits the
detection of individual microbes by hybridization analysis with specific probes, using DNA
sequencing analysis. The study of microbial activity in different habitats, for example soil,
sediments, water hydrothermal vents, mats sewage treatment plants, air etc are possible by
PCR-DGGE technique (Green et al., 2007; Liang et al., 2008). The microbial habitats in our
research are sourced from various samples of a hemicellulose-enriched sugar mill area.
2.4.3 Identification of unknown microorganisms based on 16S rDNA sequence
analysis
Small ribosomal subunits in prokaryotes contain 16S rDNA and the large subunit contains 5S and
23S rDNAs. Usually, microbial 16S, 23S, and 5S rDNA genes are arranged as a co-transcribed
operon. There are between one and several copies of the operon distributed in the genome. For
instance Ecoli has seven (Yusupov et al., 2001). The length of 16S rDNA is about 1.542
nucleotides, which are highly conserved. The 16S rDNA analysis has become a very valuable
tool, to permit identification the unknown and known microorganisms. One of the important
issues is the recognition of difficult-to-culture organisms in the laboratory from extreme
environments. Only about 1% of the community is revealed by culturing techniques. The ability
to map sequences has rapidly moved research in a different direction, towards analysis of the
hypervariabale regions of 16S rDNA. Generally these are at the edge of highly protected regions.
Primers can be designed to match with these regions and to amplify variable regions by PCR. It is
possible to identify a large number of species through DNA sequencing of 16S rDNA (Schuller et
al., 2010).
rRNA is naturally present in high copy numbers (up to 10,000 molecules per cell), it provides a
target for a highly sensitive PCR assay. rRNA molecules form a part of all ribosomes and can
therefore be used as a PCR target independent of gene expression. Computer alignment studies
of these rRNA sequences have revealed the existence of regions with highly conserved
sequences and regions which display sequence variability at the genus and species levels,
allowing the selection of genus- and species specific primers for the PCR (Daley et al., 2008).
The rDNA is the least variable gene in bacterial cells. Consequently 16S rDNA encoding genes
may be used to study the taxonomy, phylogeny and species diversity in all bacteria. So, the study
of 16S rDNA sequences reveals the genetic relatedness of microorganisms. Woese et al. (1990),
proposed the novel three domain system of categorization on the basis of such sequence data.
2.4.4 Metagenomic methods - PhyloChip

Chapter 2 Literature Review
35
DNA microarray technology offers the possibility of analysing microbial communities without
pre-requisite cultivation, thus being ideal for biodiversity studies. A DNA PhyloChip has been
developed to assess microbial diversity, based on the transfer of 16S rRNA probes from dot-blot
or fluorescent in situ hybridization (FISH) analyses to a microarray format. Similarly, it is possible
to use 16S rRNA probes to determine a signal on the microarray by the fragmentation of the 16S
rRNA molecule, or PCR amplicon, which have to be less than 150 bp in length to minimize the
formation of secondary structures in the molecules so that the probe can bind to the target site.
Liles et al., (2010) used microarray to probe for sequences based upon a phylogenetic analysis of
16S rRNA genes recovered from members of the bacterial (Acidobacteria) division. They found
that a phylogenetic microarray was useful in revealing changes in microbial population-level
distributions in a complex microbial community (Liles et al., 2010). Phylogenetic microarrays, or
“PhyloChips”, have been inceasingly used to discriminate rapidly between diverse 16S rRNA
genes present in cultured microorganisms or environmentalsamples. Compared to the labour
and resource intensive efforts to clone and sequence a representative number of clones from a
16S rRNA gene clone library, phylogenetic microarrays can provide a rapid and efficient readout
of the phylogenetic diversity present in an environmental sample. Furthermore, a hierarchical
design permits probing for microbial taxa at different phylogenetic levels (Huyghe et al., 2008),
providing information regarding the presence or absence of the many components of the tree of
life.
In general, the same strategies for in-silico development and technical set-up for fabrication and
hybridization of the RHC-PhyloChip were used as for the development of a 16S rRNA-targeted
oligonucleotide microarray for detection of all lineages of recognized sulfate-reducing
prokaryotes (SRP-PhyloChip). In addition, due to their importance for bioremediation and
agriculture, several approaches for the detection of members of the order “Rhodocyclales” have
been developed. Besides traditional cultivation methods, molecular detection of members of
this order has been based on taxon- or clone-selective 16S rRNA gene-targeted PCR primers or
probes (Loy et al., 2005). While these molecular methods were well suited for the detection of a
few selected subgroups or species within the Order Rhodocyclales, tools for surveying the
diversity of members of this Order in parallel were lacking at the time. DNA microarrays, which
have recently been introduced to microbial ecology, generally fulfilled all requirements for the
high-resolution monitoring of complex microbial communities. Their main drawback is the
extremely high cost of purchase and interpretation of the PhyloChips.
The 16S rDNA gene has been used to identify the effects on microbial community structure using
such methods as terminal restriction fragment length polymorphism (T-RFLP), length

Chapter 2 Literature Review
36
heterogeneity polymerase chain reaction (LH-PCR), denaturing gradient gel electrophoresis
(DGGE), and cloning and sequencing (Connon et al., 2005; Freeborn et al., 2005). However,
unlike these molecular analysis techniques, microarrays not only indicate general changes in
microbial community structure, but can also identify organisms present in a given sample. Our
research represents an original approach to establish better an understanding of the richness
and variety of the pentose-utilizing microbial population using a metagenomic PhyloChip
method.
This study investigates the metabolic and anabolic chanracteristics of pentose utilizing
microorganism which has not done previously. The research gaps in the literacure have
discussed in this chapter. The role of actinomycetes in sugar waste water treatment ponds and
non-sugar treatment ponds will be analysed in this research project. In addition, this research
also reports the results of analysis of the microbial diversity of microorganisms in hemicellulose-
enriched habitats.

Chapter 3 Identification
37
CHAPTER 3
Isolation and Identification of Environmental Pentose-
Utilizing Bacteria
3.0 Summary
The isolation and identification of environmental pentose-utilizing bacteria (E-PUB) are reported
in this chapter. The selected sources for this investigation were those associated with sugar-
cane mills, as the milling process results in large quantities of hemicellulose-enriched wastes.
The samples were selectively enriched in media containing single pentose sugars as the sole
carbon source. Following basic identification using phenotypic characteristics, the six remaining
isolates were identified further, using 16S rDNA testing followed by DNA sequencing. All of the
isolates were identified as members of the Order Actinomycetales, known to contain genera and
species capable of breaking down plant material, thereby producing industrially useful end
products.
3.1 Introduction
The bioconversion of pentoses derived from hemicellulose remains a problem in developing
industrial level production, as many microbes either preferentially use glucose in the presence of
mixtures of carbohydrates, or cannot use pentose sugars at all. As an initial approach towards
resolving this issue, microorganisms indigenous to a pentose-rich environment were targeted,
which are capable of metabolizing pentose sugars such as xylose, arabinose and ribose, as found
in lignocellulose.
Microorganisms such as the yeast, Rhodotorula glutinis, and bacteria such as Mycobacterium
species, Corynebacterium spp, and Clostridium acetobutylicum, are able to utilize pentoses
naturally (Singh and Mishra, 1995). A number of other naturally occurring organisms that
convert pentoses to ethanol were also named by Singh and Mishra in 1995. These included the
yeasts, Pichia stipitis, Pachysolen tanophilus and, later, Candida shehatae, which was noted by
Kastner et al., (1999).
The screening of environmental samples taken from diverse ecological niches of Queensland
sugar mills was carried out in order to explore the potential of the diversity of microbial flora of
this hemicellulose-enriched habitat. This study was undertaken with the aim of discovering the
presence of pentose-capable species of genera such as Corynebacterium and Nocardia,

Chapter 3 Identification
38
members of the class Actinomycetes. These are known to be environmental organisms with a
capacity for production of useful bioactive metabolites. These source areas are poorly studied
and may represent diverse and largely unscreened pentose utilizing capability.
3.2 Materials and Methods
3.2.1 Sample collection
This research was carried out in order to discover pentose-utilizing bacteria from soil samples
obtained from 3 different sugar mills in the State of Queensland, Australia (Figure 3.1). The
Proserpine and Maryborough sugar mills were the targeted sampling sites.
PROSERPINE
MACKAY
MARYBOROUGH
Figure 3.1: Map of the Queensland sugar mill regions
(Johnston, http://www.johnston-independent.com/ sugar.html)
Soils from areas surrounding sugar mill waste ponds were collected from the Maryborough and
Proserpine sugar mills. Each soil sample was aseptically collected into sterile test tubes and kept
at 4⁰C until they were processed in order to prevent overgrowth by fast-growing fungi and
bacteria. Processing occurred up to 2 days after sample collection. Sample details and the
collection location are detailed in Table 3.1.
ATCC cultures (Corynebacterium cystitidis (ATCC #29593) Nocardia vaccinii (ATCC #11092)) were
used as controls when required for genotypic and phenotypic comparison with isolates.

Chapter 3 Identification
39
Table 3.1: Locations of Collected Samples
Sample code Sampling location
58 Effluent Pond (58EP) Proserpine
31 Tailing Gully (31TG) Proserpine
41 Scodellaro (41S) Proserpine
46 Blair (46B) Proserpine
42 Caswell (42C) Proserpine
55 Tropic Isle (55TC) Proserpine
Batch one
Pond cell-1 (PC1) Maryborough
Pond cell-2 (PC2) Maryborough
Pond cell-3 (PC3) Maryborough
Pond cell-4 (PC4) Maryborough
Pond cell-5 (PC5) Maryborough
Pond cell-6 (PC6) Maryborough
Pond cell-7 (PC7) Maryborough
Pond cell-8 (PC8) Maryborough
Batch two
cell-1 (NC1) Maryborough
cell-2 (NC2) Maryborough
cell-3 (NC3) Maryborough
cell-4 (NC4) Maryborough
cell-5 (NC5) Maryborough
cell-6 (NC6) Maryborough
cell-7 (NC7) Maryborough
cell-8 (NC8) Maryborough
3.2.2 Isolation and Identification (culture-based)
Bacterial strains were isolated from soil samples by means of a series of enrichment steps, in
which cultures were initially inoculated with 10% wet weight of soil using Luria Bertani (LB)
(Atlas, 2004) broths containing one of 0.5% xylose or arabinose or ribose (100 mL cultures in
500 mL Erlenmeyer flasks).

Chapter 3 Identification
40
Medium was selected from the Handbook of Microbiological Media (Atlas, 2004). The isolation
medium used was minimal Luria Bertani broth with one of the pentoses provided as a sole
carbon source. This enabled a broad range of bacteria to be isolated, including Actinomycetes.
The widely used medium known as Luria Bertani broth is popular with bacteriologists because it
permits fast growth and good yields for many non-fastidious species. The medium consists of
tryptone, yeast extract and salt, adjusted to pH 7.0. While this is not strictly a “minimal medium”
such as would normally be provided for nutritional studies such as these, the intended target
organisms required a somewhat richer source of nutrient than would be supplied by a basal
medium.
To test the carbon source utilization, carbon sources such as pentoses (xylose, arabinose or
ribose), or a hexose (glucose) sugar were provided. Strains were isolated on the basis of their
carbohydrate utilization patterns, specifically growth on several types of pentoses, in the
presence and absence of glucose using LB broth, with the sugars added from sterile stock
solutions to provide the required carbon source(s). It was clear that the organisms isolated from
the environmental samples were not capable of using yeast extract as a carbon source, as a large
number of them could not grow in the broths containing pentose sugars. Had they been able to
utilize yeast extract as an alternative carbon source, these organisms would have grown in all of
the broths, regardless of the sugar provided, since yeast extract was present in all of the media.
Serial sub-culturing was undertaken following an initial 48 hours of aerated growth at 30°C and
this was repeated twice using a 1% inoculum into fresh LB broth with an antifungal agent
(nystatin powder 0.5mg/100ml) added each time. Cultures were aerated by shaking at 100 rpm,
because the target bacterial group was primarily the actinomycetes, which are all aerobes.
Subsequently, individual strains were isolated by plating onto LB agar media containing the same
set of pentoses. Isolates were stored in 50% glycerol at -20°C, prior to testing for their
carbohydrate utilization patterns (see Chapter 4).
As it was expected that suitable organisms for the purposes of this study would be those in the
class Actinomycetes, such as Nocardiaand Corynebacterium, particularly environmental species
of these genera, the following ATCC cultures were used as positive controls:
Nocardia vaccinii (ATCC #11092)
Corynebacterium cystitidis (ATCC #29593)

Chapter 3 Identification
41
Gram staining, acid fast staining, motility testing, catalase testing and spore staining were
carried out to further identify the target microorganisms according to the phenotypic
characteristics noted in Bergey’s Manual (Holt, 2000).
3.2.3 DNA analysis methods
3.2.3.1 Primer design
DNA analysis was performed in order to confirm the identity of the isolated species. Allele
specific primers for Corynebacterium cystitidis (Primer A) and Nocardia vaccinii (Primer B) were
designed using the Primer Express 2.0 primer design software programme (Applied BioSystems,
USA) and are shown below. Allele specificity is determined by the type of nucleotide base at the
3’ end of the primer sequence.Primers were synthesised by Sigma–Aldrich, Castle Hill, NSW, and
Australia.
Primer A:
5’-GAA-CGC-TGG-CGG-CGT-GCT-3’
5’-ACC-TTG-TTA-CGA-CTT-CGT-3’
Primer B:
5’-GTA-AAA-CGA-CGG-CCA-GGA-TGC-AAC-GCG-AAG-AAC-CTT-ACC-T-3’
5’-CAG-GAA-ACA-GCT-ATG-ACT-ACG-GCT-ACC-TTG-TTA-CGA-CTT-CG-3’
3.2.3.2 Extraction of DNA
DNA was extracted using the QIAGEN DNeasy kit 2006 according to the instructions for Gram-
positive bacteria. The DNeasy Mini spin column was used. DNA was then eluted with buffer for
DNA purification. Presence of DNA was tested using absorbance ratio (A260:280). The critierion
for acceptability range was 1.8 to 2 and all of the results fell into this range.
3.2.3.3 PCR amplification of 16S rDNA
The reaction mixture contained master mix with water, buffer, dNTPs, primers and Taq DNA
polymerase in a single tube. The mixture was then aliquoted into individual tubes. MgCl2 and
template DNA solutions were added to the micro-tube (reagent details Table 3.2), which was
then placed on a thermocycler (BioRad, DNA engine, Peltier, Thermal cycler) to start the PCR
reaction (Bio-Rad laboratories 2008).

Chapter 3 Identification
42
Table 3.2: PCR reaction mixture details
Reagent Final Concentration Quantity for 50 µL of
reaction mixture
Sterile deionized water - Variable
10X MgCl2 buffer 1X 3.5 µL
2 mM dNTP mix 0.2 mM of each 2.5 µL
Forward Primer 1 µM 1 µL
Reverse Primer 1 µM 1 µL
Taq DNA Polymerase 1.25 u / 50 µL 0.2 µL
Template DNA 1 µg 1 µL
Reactions were carried out in a thermal cycler under the following conditions: initial
denaturation at 94 °C for 3 min; 40 cycles of 95 °C for 1 min, 55 °C for 40 s and 70°C for 1 min 30
sec; final extension at 70 °C for 5 min.
3.2.3.4 Gel electrophoresis
To prepare the gel, a 2% agarose (2% agarose gel on TAE buffer) solution was heated (65 C -
85 C) in a microwave ovenfor about 2 min and then cooled to about 55°C, the gel poured and
the comb inserted. Once solid the gel was covered with 1X TAE running buffer and 5 µL of DNA
mixture was loaded into each well. Samples were run with 80-100 volts for 30-45 minutes.
Finally the gel was washed with buffer (0.5% ethidium bromide: 200 mL 1X TAE) and visualized
under UV light.
3.2.3.5 16S rRNA gene sequencing
The 16S rRNA was sequenced using a Big Dye Terminator Cycle Sequencing reaction. The 20-μL
sequencing reaction consisted of 1 μL Big dye, 3.5 μL sequence buffer 12.5 μL of water, 3.2 pmol
of sequencing primer, and 200 ng PCR product. Cycle sequencing was performed using a Gene
Amp PCR system (BIO-RAD, DNA engine, Peltier, Thermal cycler), programmed for 30 cycles at
96°C for 20sec, 50°C for 20sec, and 60°C for 4 min using 16S forward and reverse primer
sequences A and B from Section 3.2.2.1. Sequencing products were cleaned with 70% ethanol
wash, using ethanol/EDTA methods according to the Griffith University sequencing preparation
instructions. Sequence products were analysed using an ABI 3500 Sequencer (Applied
Biosystems). 16S rDNA sequences were assembled from forward and reverse analyses and then

Chapter 3 Identification
43
edited with Sequencer software (Bio- Edit). 16S rDNA sequences results were compared with
sequences in GenBank, using the BLAST sequence similarity search.
3.3 Results
3.3.1 Primary identification of isolates
This research conducted all series of experiments in duplicate, to identify the target isolates. The
aim was to isolate actinomycetes, and similar or related organisms. Six cultures proved to be of
great interest from 191 isolates, from eleven different environmental sources (six samples from
Proserpine, and five from Maryborough). The characteristics of these isolates are summarized in
Tables 3.3, 3.4 and 3.5.
There were two different colony types found from the Maryborough mill (A) samples, similar to
the expected appearance of Corynebacterium, Nocardia and Mycobacterium on the basis of
colony descriptions in Bergey’s Manual (Holt, 2000). Colonies found in samples from both
Maryborough mill samples (A and B) and Proserpine mill, are described in Tables 3.3, 3.4 and 3.5
respectively. It should be noted that many of these colony types were very similar in
appearance. The biochemical tests matched correctly with the properties expected of
Actinomycetes (Tables 3.3, 3.4 and 3.5).
Table 3.3: Characteristics of selected isolates from Maryborough (A)
Sample
source
Colony appearance Biochemical properties
Maryborough
(A)
LB(0.5%
xylose)
LB(0.5%
arabinose)
LB(0.5%
ribose)
Gram
stain
Acid
fast
Catalase
test
Motility
test
Endospore
stain
PC1-2
slow growing
white, big dry
colony
_
_
+ rod
_
+
_
_
PC4-1 cream,
irregular,
filamentous
cream,
irregular,
filamentous
_
+ rod
_
+
_
_

Chapter 3 Identification
44
Table 3.4: Characteristics of selected isolates from Maryborough (B)
Sample
source
Colony appearance Biochemical properties
Maryborough
(B)
LB(0.5%
xylose)
LB(0.5%
arabinose)
LB(0.5%
ribose)
Gram
stain
Acid
fast
Catalase
test
Motility
test
Endospore
stain
NC1-3
cream,
irregular, flat,
smooth
rhizoid
surface
cream,
irregular, flat,
smooth
rhizoid
surface
_ + rod _ + _ _
NC1-2 white
creamy,
irregular, flat
surface,
_ _ + rod _ + _ _
NC4-1 cream,
irregular,
filamentous
cream,
irregular,
filamentous
cream,
irregular,
filamentous
+ rod _ + _ _
Table 3.5: Characteristics of selected isolates from soil of Proserpine
3.3.2 16S rDNA analysis
PCR analysis of 16S rDNA was performed to confirm the species level identification of the E-PUB
microbes. In Figure 3.2 a, the lanes 2-7 inclusive contained the isolates NC1-2 NC1-3, PC1-2, PC4-
1, NC4-1, 31TG-3, which were amplified with A primer.
Sample
source
Colony appearance Biochemical properties
Proserpine LB(0.5%
xylose)
LB(0.5%
arabinose)
LB(0.5%
ribose)
Gram
stain
Acid
fast
Catalase
test
Motility
test
Endospore
stain
31TG-3 _ Light cream,
smooth, very
small in size
_ +
cocco
bacilli
_ + _ _

Chapter 3 Identification
45
a. PCR amplification with primer A (6 unknown isolates)
Figure 3.2: 16S rDNA PCR amplification image after gel electrophoresis
Primer B effectively amplified the isolates NC1-2 NC1-3, PC1-2, PC4-1, NC4-1, 31TG-3, as shown
on Figure 3.2b. The DNA of all indigenous microbes displayed bands on agarose gel after
amplification with primers A and B, which were designed to detect the Order Actinomycetes.
Primer A was designed to detect bacteria similar or identical to Nocardia species, and Primer B
PC1-2 PC4-1 NC1-3 NC1-2 NC4-1 31TG
Indicating 5000 BP according to the marker
Amplification with primer A
PC1-2 PC4-1 NC1-3 NC1-2 NC4-1 31TG
PC1-2 PC4-1 NC1-3 NC1-2 NC4-1
Amplification with primer A 5000 BP
Amplification with primer B and 3000 BP
b. PCR amplification with primers A and B

Chapter 3 Identification
46
was designed to identify coryneform types and organisms closely related to the
Corynebacterium.
3.3.3 16S rDNA gene sequencing
The sequences were compared using the BLAST program with the sequences in the NCBI
database. Using a threshold of greater than 77% similarity for positive identification, evidence
was found for the presence of Corynebacterium spp., Actinomycess spp., Rhodococcus spp.,
Propionibacterium spp., and Nocardia sp (Table 3.6). Twenty four (both forward A, B and reverse
A, B) sequences were obtained, only partially overlapped with the sequences from Genbank
because the sets of primers (forward and reverse) A and B were used for sequencing for
database entries. Primer A was designed to detect relatives of Corynebacterium and Primer B
was designed for Nocardia-like species. The sequences were aligned according to the similarity
between these sequences and taxonomically placed with microorganisms in Genbank. The list of
identified bacteria found from the pentose-enriched habitat is given in Table 3.6.
Table 3.6: 16S rDNA sequencing results of unknown isolates
The sequencing results, within the limitations of primers designed on partial sequences, were as
expected. They demonstrated that the isolates were all known soil organisms and members of
the Order Actinomycetales.
Isolate
code
Closest matched
Genus/Species by
Primer A
Sequence
similarity
in % (no.
of bases)
Taxonomic
group
Closest matched
Genus/Species by
Primer B
Sequence
similarity
in % (no.
of bases)
Taxonomic
group
PC1-2 Corynebacterium glutamicum
98% Actinobacteria
Corynebacterium freiburgense
80% Actinobacteria
PC4-1 Actinomyces odontolyticus
77% Actinobacteria
Actinomyces odontolyticus
83% Actinobacteria
NC1-3 Actinomyces odontolyticus
77% Actinobacteria
Actinomyces odontolyticus
83% Actinobacteria
NC1-2 Corynebacterium glutamicum
98% Actinobacteria
Corynebacterium freiburgense
80% Actinobacteria
NC4-1 Rhodococcus equi 98% Actinobacteria
Nocardia elegans 87% Actinobacteria
31TG Propionibacterium freudenreichii
79% Actinobacteria
Propionibacterium freudenreichii
99% Actinobacteria

Chapter 3 Identification
47
3.4 Discussion
Six indigenous cultures belonging to the Order Actinomycetales were isolated from soil samples
of sugar mill treatment ponds in Proserpine and Maryborough sugar mills. The strains were
tagged as NC1-2, NC1-3, PC1-2, PC4-1, NC4-1, 31TG-3 and showed typical Actinomycete
phenotypic (physical and biochemical) characteristics such as a typical rhizoid colony edge,
Gram-positivity, and catalase-positivity.
The analysis of the complete or partial (500 bp of the 5′ end) 16S rRNA gene is the most
extensively used molecular method for species recognition and has a broad database for
comparison (Daley et al., 2008). Under routine conditions, the method is useful and convenient.
The 16S rDNA analysis identified the six indigenous isolates as belonging to four different species
of microorganism. The 16S rDNA sequencing confirmed that primers A and B matched regarding
the identity of two of the isolates: PC4-1 and NC3-1 were both identified as Actinomycetes
odontolyticus; and 31TG was identified as Propionibacterium freudenreichii. Two isolates were
identified as Corynebacterium genus, with the species being either C. freiburgense or C.
glutamicum (PC1-2 and NC1-2). Obviously these species are taxonomically close. The final
isolate, NC4-1 was identified as either Rhodococcus equi or as Nocardia elegans.
Previous research surveys indicate that Actinomyces, Corynebacterium, Nocardia, and the
Rhodococcus complex form distinct taxa of equivalent rank (Embley and Stackebrandt, 1994; Liu
et al., 1996). Propionibacterium is separated from Nocardia according to the oxygen
requirement for growth. Propionibacterium is a facultative aerobe and Nocardia is strictly
aerobic. Nocardia is related to Corynebacterium and Rhodococcus in terms of fatty acid
sequences in the cell wall. Bergey’s Manual (Holt,2000) suggest that Rhodococcus, Nocardia and
Corynebacterum are members of the same family. According to the scientific classification, all of
the isolated bacteria belong to the same Order, the Actinomycetales (Embley and Stackebrandt,
1994; Liu et al., 1996).
Rhodococcus has had several taxonomic designations in recent years (Tsukamura, 1982;
Goodfellow et al., 1998), and the genus most frequently confused with Rhodococcus is Nocardia.
It is therefore evident that these genera are very similar, and according to the identity criteria, it
is not surprising that they have both been associated with the same isolate (refer to Table 3.6:
Result of 16S sequencing). Furthermore the phylogenetic relationships between Rhodococcus,
Mycobacterium, Nocardia and Streptomyces are very close, and data from Mordarski et al.,
(1980) represented the genera Rhodococcus, Mycobacterium, Nocardia and Streptomyces as

Chapter 3 Identification
48
four recognizable clusters on the similarity map. Rhodococcus equi was poorly defined compared
to Nocardia elegans on BLAST analysis of 16S sequencing.
It is mentioned in Bergey’s Manual (Holt, 2000) that the genus Propionibacterium is also very
frequently confused with Corynebacterium.
Two isolates were identified as being either C. glutamicum or C. freiburgense. The main
difference between species is that C. glutamicum is a non-pathogenic bacterium. On the other
hand, C. freiburgense is pathogenic and is a very recently identified species (Funke et al., 2009).
The characteristics of C. freiburgense are not yet included in Bergey’s Manual. The distinctive
“wagon-wheel” colony appearance (beige–whitish, dryish, convoluted with irregular edges)
described for C. freiburgense (Funke et al., 2009) was not observed in our isolates, so it is
considered most likely that the isolates were C. glutamicum.
It was also mentioned in Holt (2000) that, the actinomycetes showed variable catalase reactions,
however, the Actinomyces spp. isolated in this research were all catalase positive. Previous
research has shown that Corynebacterium, Nocardia and Rhodococcus (Order Actinomycetales)
are mostly pathogenic (Barry and Beaman, 2006) but our isolates were not clinical, being found
instead in the common source of environmental soil. According to Bergey’s Manual (Holt, 2000)
and to Laurent et al., (1999) most of the species of Corynebacterium, Nocardia and Rhodococcus
are widely distributed but particularly abundant in soil which supports our identification of these
isolates as being in this group
Pentose-degrading microorganisms are relatively rare in natural environments and agricultural
soils (Gírio et al., 2010). In the current study, six indigenous members of the Order
Actinomycetales were isolated from hemicellulose-enriched soil from a total of 191 initial
isolates. The strains were given the opportunity to induce their ability to grow in the presence of
different pentose sugars (xylose, arabinose and ribose) as carbon sources. Therefore, the use of
these wild-type microorganisms from soils is an attractive approach for industrial isolates, since
they have already adapted to the pentose-rich habitat of sugar treatment ponds. Samples from
the sugar treatment pond are expected to be hemicellulose-enriched, as sugar cane has high
levels of lignocelluloses, and pentoses are the most abundant carbon source in hemicellulose
(Patel et al., 2004). The microbes found in those habitats were therefore expected to be adapted
to utilize pentose as a carbon source of nutrient. The microbes isolated from the soil of sugar
treatment ponds proved to be common soil bacteria.
Propionibacterium freudenreichii and Actinomyces odontolyticus are potential human pathogens
whereas Rhodococcus equi and Nocardia elegans are usually animal pathogens (Lemee et al.,

Chapter 3 Identification
49
1994). There are several characteristics of Corynebacterium glutamicum that make it useful for
industrial biotechnology processes. It is not pathogenic, does not form spores, grows quickly, has
relatively few growth requirements, has no extracellular protease secretion and has a relatively
stable genome (Mateos et al., 2006; Burkovski, 2008). C. glutamicum produces several useful
compounds and enzymes. It was first discovered as a producer of glutamate. Now it is also used
to make amino acids, such as lysine, threonine, and isoleucine, as well as vitamins like
pantothenate (Kalinowski et al., 2003; Kirchner and Tauch, 2003).
Another potential use for C. glutamicum is in bioremediation, such as for the remidiation of
arsenic residues. C. glutamicum contains two operons in its genome, the ars1 and ars2 operons,
with further experimentation. Researchers hope to be able to eventually use this bacterium to
take up the arsenic in the environment (Mateos et al., 2006).
3.5 Conclusions
The bacteria isolated and identified in this research are natural isolates from soil samples rich in
hemicellulose, obtained from sugar mills in Queensland, Australia. They were not genetically
modified to promote the preferential use of pentoses. These E-PUB were able to utilize pentose
sugar as a carbon source during their growth on LB medium, following a period of
acclimatization to the supplied pentose carbon sources. It was found that each of the E-PUB
grew on different types of sugar provided as carbon sources. In the absence of a suitable added
carbon source, no growth was observed, so demonstrating that the growth was not due to the
presence of yeast extract or other media constituents.
The isolates were identified as various genera, all members of the Order Actinomycetales.
Members of this order are well known for their ability to produce bioactive metabolites and are
widely distributed in nature.
Further research will be reported in the following chapters, regarding the nature and extent of
the pentose-degradation process, and the resulting end-products.

Chapter 4 Catabolic Characteristics of E-PUB
50
CHAPTER 4
Catabolic Characteristics of Environmental Pentose-
Utilizing Bacteria (E-PUB)
4.0 Summary
In order to examine the metabolic characteristics of the microorganisms isolated from pentose-
enriched samples obtained from sugar cane mills, it was first necessary to test the capability of
those organisms to break down pentose sugars such as those found in hemicellulose. This
chapter describes catabolic characteristics of E-PUB by growing them in media of single pentose
sugar carbon sources, and subsequently in media containing glucose in addition to each pentose
sugar (dual sugar). A set of pentose sugars was selected as being representative of those found
in the natural environment under examination, i.e. the waste products of sugar cane milling. The
organisms tested, having been isolated according to their ability to use pentose sugars (reported
in Chapter 3), were all capable of growing on at least one of the pentoses provided. All were also
able to grow on mixtures of glucose with at least one pentose sugar, demonstrating a
characteristic diauxie growth pattern.
4.1 Introduction
As was discussed in Chapter 2, there are several difficulties with respect to the use of
hemicellulose as a raw material for the industrial production of commercial products. Principal
among these is the fact that the raw material contains a mixture of sugars, including both
hexose sugars, mainly glucose, and pentose sugars, such as xylose, arabinose, and ribose. Most
of the microorganisms capable of fermenting glucose to give high yields of appropriate end-
products, are incapable of also fermenting the pentose sugars, or do so at such low yields that
the proposition is not cost effective (Agbogbo et al., 2006). There has been some discussion
(Pitkänen et al., 2005; Agbogbo et al., 2006; Hahn-Hägerdal et al., 2007) regarding the best
means by which to utilize most efficiently a large component of the sugars present in the
agricultural waste material. One approach to resolving this problem has been to genetically
engineer such organisms, to introduce the additional capacity for pentose-utilization (Pitkänen
et al., 2005; Hahn-Hägerdal et al., 2007).

Chapter 4 Catabolic Characteristics of E-PUB
51
Another possibility has been the suggested use of a mixed starter culture, such that one
organism will use glucose and a second organism would breakdown the pentose substrates.
Finally, the method employed here is to isolate natural, indigenous microorganisms from a
pentose-rich environment, which may reasonably be expected to be pentose-capable, as well as
having the ability to breakdown the more common substrate, glucose. Such isolates were
discovered, as described in Chapter 3, by screening process using pentose substrates.
These isolates were grown in the test pentose sugars separately, and also with glucose, and their
specific growth rates during the exponential phases were compared. Further, their ability to use
glucose and pentose sugars as dual carbon sources was examined. The results of this research
are presented in this chapter. The aim of this research was to study the growth patterns of
microorganisms that could effectively use pentose sugars, both alone, and as a mixture with
glucose.
There were a number of ways by which the utilization of dual sugar substrates might be
accomplished. It is possible that, as the organisms were initially isolated by growth on a pentose
sugar carbon source, they could therefore be pre-induced to pentose-utilization, and were also
constitutively capable of breaking down glucose, as indeed are most microorganisms. In this
scenario, the microorganisms could be expected to use both glucose and pentose sugars
simultaneously, resulting in a rather long exponential growth phase, and a growth rate similar to
that of glucose growth conditions.
More likely, however, is the expectation that the organisms will use a diauxie growth pattern
(Brückner and Titgemeye, 2002), whereby they use glucose first, and, after a second lag phase,
launch into a second exponential growth phase, based on the pentose sugar. This growth
pattern is due to the catabolite repression initiated by the metabolism of glucose, which
prevents simultaneous utilisation of any other substrate. The presence of a diauxie growth
pattern is easily observed by measuring the growth curve of the microorganisms while they
grow in a mixed sugar environment.
In order to calculate the appropriate ratio of sugars for this research, natural bagasse was used
as a source. The partial composition of natural sugarcane bagasse is xylose 25.2 and glucose
41.0, expressed as % w/w of the dry matter (Pandey et al., 2000).

Chapter 4 Catabolic Characteristics of E-PUB
52
4.2 Materials and Methods
Bacterial culture Corynebacterium cystitidis (ATCC 29593), and isolates Propionibacterium
freudenreichii; (31TG) Corynebacterium glutamicum (PC1-2, NC1-2); Actinomyces odontolyticus
(PC4-1, NC1-3) and Nocardia elegans (NC4-1), were used in this study. Duplicate E-PUB cultures
were performed for the analysis of catabolic characteristics. The ATCC culture was used as a
positive control as it was expected that the bacterial isolates would have similar metabolic
characteristics to that culture. The bacteria were recovered from storage in -20°C glycerol stock
by growing them overnight at 30°C in 100 mL of minimal medium (Luria Bertani broth)
containing 0.016 M pentose sugars (xylose or arabinose or ribose).
The overnight cultures were diluted in fresh medium to an approximate concentration of
107cfu/mL, and subsequently incubated at 30°C for approximately 3 days. The resulting biomass
was collected by filtration onto a membrane filter (pore size = 0.2 µm), and then washed with
warmed (to avoid cold-shock on the organism) minimal medium without any carbon sources
(Wittmann et al., 2004). The isolates were then cultured in Luria Bertani (LB) broth
supplemented with 1.0% (w/v) xylose, arabinose or ribose from 20% sterile stock solutions, at
30⁰C in a shaking incubator (Ratek, Orbital Mixer Incubator) at 100 rpm.
Growth was determined by measuring the optical density at 680 nm, using a spectrophotometer
(Beckman Coulter Spectrophotometer) with measurements being taken at 12 hourly intervals
against a medium blank. Optical density was plotted against time in hours from the inoculation
time. The OD was measured at 680 nm, rather than 540 nm as is generally used for bacteria,
because the actinomycetes cells form a mass of cell-chains (Janssen et al., 2001) making the
longer wavelength more effective for measuring optical density. The optical density (OD680) was
measured after 24 hrs incubation and then at 12 hourly intervals until the stationary phase was
reached. The OD value was converted to logarithmic value. Two OD values at the beginning and
end of the exponential phase were selected for OD1 and OD2 and the corresponding times t2 and
t1 values, and used in following equation to calculate μ the specific growth rate, µ, of each
exponential phase:
Next, the specific growth rates for growth on glucose and pentose substrates were compared
using the student T test and SPSS software. The “Independent samples” t-test was used to
decide whether two means were significantly different from each other when the two samples
were taken from different values. The significant p value was considered to be below 0.05.
12
12 lnln
tt
ODOD

Chapter 4 Catabolic Characteristics of E-PUB
53
Following the single carbon source experiments, the same set of organisms was tested in the
same way, but with two sugars being present, these being glucose plus one of the pentose
sugars. The pentose sugar, xylose, is commonly found in agricultural residues and other
lignocellulosic biomasses, but is not the only sugar present. In order to evaluate whether xylose
(or other pentose sugar) catabolism by the indigenous isolates background was repressed in the
presence of glucose, a culture broth of each of the bacterial cultures (the six isolates plus the
positive ATCC control) was prepared in a mineral medium containing xylose, arabinose or ribose.
A sample from each of the cultures was subcultured into media containing each of the following
dual-sugar carbon sources: glucose-xylose; or glucose-arabinose; or glucose-ribose.
Initially, various concentrations of both glucose and the pentose sugars were tested using the
ATCC culture, to establish the optimum balance, which was found to be 50 µL each of glucose
(0.025g or 2.5%) and xylose/arabinose/ ribose (0.0125g or 1.25%) per 100 mL broth medium.
Stock solutions of carbon sources were made as: glucose 0.5 g/mL and pentoses 0.25 g per mL.
This resulted in an almost identical ratio (2:1) of hexose: pentose as that found in naturally in
hemicellulose material (1.6:1).
The broths were incubated at 30°C in a shaking incubator (Ratek: Orbital mixture Incubator) at
100 rpm and the optical density was measured at 12 hourly intervals as per the previous
experiment (above).
Triplicate glucose measurement was performed using Megazyme-Glucose assay kit. Glucose
Reagent Buffer was (25.0 mL) diluted in to 500mL distilled water. All contents of one vial Glucose
Determination Reagent were dissolved in this buffer. The absorbance of blank, standard and
sample was measured at 510 nm after mixed and incubates at 40 or 50°C for 20 min. Glucose
content was calculated by using following formula:
Calculation:
4.3 Results
4.3.1 Comparison of growth using different single carbon sources
This research consisted of a series of growth experiments to investigate the target
microorganism’s catabolic characteristics. The ability of the test organisms to grow using various
carbon sources was measured by observation of the final optical density, indicating increasing
biomass and the efficiency of this utilisation was noted by calculation and comparison of the
specific growth rates during the exponential phases of growth in each case.

Chapter 4 Catabolic Characteristics of E-PUB
54
A possible confounding factor was that the medium contained yeast extract, which could
potentially have acted as a carbon source in its own right, so negating the results of the “single
carbon source” experiments. However, the results show that, while the E-PUB grew on media
that contained different types of sugar as carbon source, some of the sugars consistently did not
support growth. From this evidence, it has can be taken that yeast extract is not sufficient to
support microbial growth as a carbon source for the tested organisms, since, if this were the
case, all of the test systems would have shown a level of background growth due to this source.
Various degrees of E-PUB growth in different pentose sugars revealed that they used the
different pentose sugars as an essential carbon source for growth.
Figure 4.1 illustrates graphically the results of the carbon source utilisation experiments,
depicting for each organism (ATCC strain and the six test isolates), the extent to which it was
able to grow on each carbon source over a period of 4 days, which was in all cases sufficient time
to reach the stationary phase.
In all cases, when the pentose sugars were able to be used, similar final optical density readings
were obtained, indicating that the pentoses were suitable as carbon sources in terms of growth
levels. In general, pentose sugars required longer lag periods than that of glucose, implying the
involvement of an inducible enzyme system, rather that the constitutive one available for
glucose metabolism. However, this was not the case for Actinomyces odontolyticus and
Corynebacterium glutamicum, so a different system may be in place for these genera.
The results clearly demonstrated that all six target isolates could utilize pentose sugars or
glucose to varying, but quite similar, extents. It was considered useful to determine the
efficiency of the metabolism (capacity to utilize carbon sources), by comparing the specific
growth rates of each species. These were calculated based on the OD measurements, and are
shown in Table 4.1. While it may be argued that specific growth rates based on OD are not
necessarily the most accurate, in the case of these organisms, with their “tangled mat” growth
habit, the OD is considered to be equally as accurate as most other commonly used measures of
bacterial growth. C. glutamicum (PC1-2) and C. glutamicum (NC1-2) had different growth rates
in terms of xylose (0.016 and 0.006) and glucose (0.022 and 0.007) utilization even though they
are the same species albeit from different sources. It may be that they are different strains of
the same species. N. elegans (NC4-1) was the only one among E-PUB was able to utilize ribose
(µ=0.009).

Chapter 4 Catabolic Characteristics of E-PUB
55
a b
c d
e f
g
X= xylose, A= arabinose, R= ribose and G= glucose, LB=
Luria Bertani (Appendix Table B1, B3, B5, B7, B9, B11,
B13).
Figure 4.1 (a-g): Comparison of growth curves for various carbon sources. Cultures included an
ATCC culture and isolates from sugar treatment ponds; optical density at 680nm was measured
against time.
0
0.5
1
1.5
2
2.5
24 36 48 60 72 84 96
OD
680
TIME
Corynebacteria cystitidis
LBX
LBG
0
0.5
1
1.5
2
2.5
24 36 48 60 72 84 96
OD
680
TIME
31TG (P. freudenreichii)
LBA
LBG
0
0.5
1
1.5
2
2.5
72 96 120 144 168
OD
680
TIME
PC4-1 (A. odontolyticus)
LBXLBALBG
0
0.5
1
1.5
2
2.5
3
0 12 24 36 48 60 72 84 96
OD
680
TIME
PC1-2 (C.glutamicum)
LBX
LBG
0
0.5
1
1.5
2
2.5
36 48 60 72 84 96
OD
680
TIME
NC1-3 (A.odontolyticus)
LBX
LBA
LBG
0
0.5
1
1.5
2
2.5
72 96 120 144 168
OD
680
TIME
NC1-2 (C.glutamicum)
LBX
LBG
0
0.5
1
1.5
2
2.5
72 96 120 144 168
OD
680
TIME
NC4-1 (N. elegans)
LBX
LBA
LBR
LBG

Chapter 4 Catabolic Characteristics of E-PUB
56
Table 4.1: Specific growth rates of E-PUB cultures. Cultures included control ATCC culture and
known isolates, using single carbon source - pentose sugars or glucose - as carbon sources.
These results showed that the six indigenous isolates, PC4-1 and NC1-3 (A. odontolyticus), PC1-2
and NC1-2 (C. glutamicum), NC4-1 (N. elegans) and 31TG (P. freudenreichii), could utilize
pentose and glucose with approximately the same efficiency as indicated by the very small
difference in specific growth rates. Such differences may, in fact, be a result of the slight
inaccuracy of using OD to calculate the growth rates.
The specific growth rates of all organisms using pentose sugars (mainly xylose) and glucose were
compared by using paired t- test in Statistical Package for the Social Sciences (SPSS) software.
Statistically significant difference would be indicated by a p value of <0.05. A statistically
significant difference was found (Table 4.2) between xylose and glucose utilization by all of the
environmental isolates and the ATCC control, even though the actual growth rates appeared to
vary, as the p value of the comparisons between specific growth rates was less than 0.05.
Specific growth rate (h−1)
Pentoses Hexoses
Bacteria Xylose Arabinose Ribose Glucose
Corynebacteria cystitidis 0.018 - - 0.016
A. odontolyticus (PC4-1) 0.012 0.007 - 0.009
C. glutamicum (PC1-2) 0.016 - - 0.022
C. glutamicum (NC1-2) 0.006 - - 0.007
A. odontolyticus (NC1-3) 0.022 0.013 0.009
N. elegans (NC4-1) 0.022 0.009 0.009 0.021
P. freudenreichii (31TG) - 0.005 - 0.011

Chapter 4 Catabolic Characteristics of E-PUB
57
Table 4.2: Statistical analysis of p values of specific growth rates. Comparison of µ using a
pentose sugar compared to that of glucose
* Arabinose specific growth rate included, as this isolate does not use xylose
4.3.2 Bacterial growth using combination of pentose and hexose (dual carbon sources) sugars
The same set of cultures as those that had been tested in the previous section, was again tested
using similar preparation methods, but this time using a glucose plus pentose dual carbon source
combination.
During the initial cultivation on single pentose sugar substrates, it was seen that the bacterial
isolates were able to utilize the pentose sugars, and were therefore using pentose metabolic
pathways. This may have led us to expect pentose metabolism to begin without a lag phase.
However, cultivation on the dual sugars (Figure 4.2 a, b, c) showed that all isolates preferred to
utilize glucose first and then they entered a stationary/lag phase following glucose depletion,
before the pentose-stimulated growth phase.
Bacteria p value (µ for
xylose vs glucose)
Corynebacteria cystitidis 0.047
A. odontolyticus (PC4-1) 0.040
C. glutamicum (PC1-2) 0.041
C. glutamicum (NC1-2) 0.048
A. odontolyticus (NC1-3) 0.03
N. elegans (NC4-1) 0.047
P. freudenreichii (31TG)* 0.03

Chapter 4 Catabolic Characteristics of E-PUB
58
a
b
c
Figure 4.2: Results of the growth measurements (OD680) for the various cultures.
Graphs depict one sugar combination per graph (a, b and c), and show growth patterns of the six
cultures tested, which were capable of utilizing the dual-sugar carbon source, on each graph
(Appendix Tables B15, B16, B17).
0
0.5
1
1.5
2
2.5
3
0 12 24 36 48 60 72 84 96 108
OD
time
glu+xyl
PC1-2PC4-1NC1-2NC1-3NC4-1
0
0.5
1
1.5
2
2.5
3
24 36 48 60 72 84 96
OD
time
glu+ara
PC4-1
NC4-1
NC1-3
31TG-3
0
0.5
1
1.5
2
2.5
0 12 24 36 48 60 72 84 96 108
OD
time
glu + rib
NC4-1

Chapter 4 Catabolic Characteristics of E-PUB
59
This second lag phase, observed before the pentose sugar exponential phase, was present
despite pre-enrichment of the bacteria with the pentose sugar, indicating that catabolite
repression was indeed taking place, inhibiting the utilization of the pentose until the hexose was
exhausted which was proven by residual glucose measurement (Table 4.5).
C. glutamicum (PC1-2, NC1-2); A. odontolyticus (PC4-1, NC1-3) and N. elegans (NC4-1) isolates
showed diauxie characteristics with glucose/xylose culture media (Figure 4.2 a). However, only
N. elegans (NC4-1) was able to utilize ribose. On the other hand, A. odontolyticus (PC4-1, NC1-
3)and N. elegans (NC4-1) and P. freudenreichii (31TG) indigenous isolates were able to utilize
arabinose after glucose and demonstrated diauxie growth characteristics.
The diauxie experiments demonstrated that all of the organisms could use a hexose and pentose
sugar sequentially. However, only one isolate, N. elegans (NC4-1) metabolised the glucose and
ribose combination, resulting in the same diauxie effect as the other isolates and sugar
combinations. As in the previous single sugar experiments, the specific growth rate for each
exponential phase of growth was calculated, to determine the efficiency of sugar utilization.
Table 4.3: Analysis of diauxie specific growth rates
Specific growth rates (h−1)
Dual sugar carbon source Glucose/Xylose Glucose/Arabinose Glucose/Ribose
Bacteria Glu Xyl Glu Ara Glu Rib
C. glutamicum (PC1-2) 0.050 0.010 - -
C. glutamicum (NC1-2) 0.013 0.008 - -
A. odontolyticus (PC4-1) 0.031 0.006 0.051 0.005
A. odontolyticus (NC1-3) 0.008 0.007 0.051 0.007
N. elegans (NC4-1) 0.032 0.008 0.055 0.004 0.04 0.006
P. freudenreichii (31TG) 0.052 0.004

Chapter 4 Catabolic Characteristics of E-PUB
60
The specific growth rate results from Table 4.3 indicate, as would reasonably be expected, that
the µ value for the glucose component of the growth curves remained reasonably consistant,
demonstrating that the presence of the pentose sugar did not dramatically affect the rate of
glucose utilization. In all cases, a lag period was observed between the glucose and pentose
growth curves, defining the diauxie phenomenon. In the case of xylose and ribose, the lag period
was generally approximately 24 hours; however the arabinose growth periods began only 12
hours after completion of the glucose curves. It is not known whether this lag period is
important, but for practical reasons, it may be useful for industrial purposes to be aware of such
a pause in the metabolic processes.
Table 4.4: P values of significant difference between specific growth rates of dual-sugar
combinations of glucose and pentose sugars
The p values of specific growth rate for glucose and pentoses utilization are presented in Table
4.4. All p values showed no significant difference between specific growth rates for hexose and
pentose-utilization as the p values are below 0.05. The sole exception is N. elegans (NC4-1)
which had p value 0.13 means indicating a significant difference between the rates of growth
using glucose as opposed to arabinose as a carbon source combination.
Residual glucose measurement provided information about the extent of glucose depletion at
the time of second lag phase.
Sugar
combination
C.
glutamicum
( PC1-2)
C.
glutamicum
( NC1-2)
A.
odontolyticus
( PC4-1 )
A.
odontolyticus
( NC1-3)
N.
elegans
( NC4-1)
P.
freudenreichii
(31TG)
glucose and
xylose
combination
0.004
0.011
0.01
0.001
0.001
glucose and
arabinose
combination
0.01 0.03 0.12 0.04
glucose and
ribose
combination
0.035

Chapter 4 Catabolic Characteristics of E-PUB
61
Table 4.5: Amount of glucose at the second lag phase of the growth medium
* The starting concentration of glucose was 2.5% (w/v)
All of the E-PUB organisms utilized glucose during the first log phase in slightly varying amounts
as shown in Table 4.5. Corynebacterium glutamicum used the most glucose, prior to using
xylose, while Nocardia elegans used the least glucose of all tested, when in combination with
ribose, although the shorter incubation time may have influenced this figure.
Table 4.6: Ratio of specific growth rate (µ) for xylose utilization as a single sugar compared to
that of the same sugar as part of a dual carbon source system.
Culture µ for Single sugar (µs) µ for Dual sugar (µd) Ratio µs: µd
C. glutamicum (PC1-2) 0.018 0.010 1.9
C. glutamicum (NC1-2) 0.022 0.008 2.75
A. odontolyticus (PC4-1) 0.010 0.006 1.61
A. odontolyticus (NC1-3) 0.009 0.007 1.14
N. elegans (NC4-1) 0.006 0.008 0.07
P. freudenreichii (31TG) 0.009 (arabinose) 0.015 0.60
Bacteria Residual amount of glucose* (g/100mL) Time
(h)
Glucose/Xylose Glucose/Arabinose Glucose/Ribose
C. glutamicum (PC1-2) 0.0040 or 0.4% 60
C. glutamicum (NC1-2) 0.0037 or 0.3% 60
A. odontolyticus (PC4-1) 0.0013 or 0.1% 0.0009 or 0.09% 60
A. odontolyticus (NC1-3) 0.0018 or 0.1% 0.0005 or 0.05% 60
N. elegans (NC4-1) 0.0002 or 0.2% 0.0041 or 0.4% 0.0001 or
0.01%
48
P. freudenreichii (31TG) 0.0006 or 0.06% 60

Chapter 4 Catabolic Characteristics of E-PUB
62
It is apparent from Table 4.6 that xylose was used more efficiently as a single carbon source than
as part of a dual carbon source system by C. glutamicum and A. odontolyticus, while both N.
elegans and P. freudenreichii were somewhat more efficient in breaking down xylose following
glucose metabolism than as a sole carbon source, although the difference was not statistically
significant. The latter two isolates are therefore better choices for industrial purposes than the
former pair, providing other qualities of the cultures are similar.
4.4 Discussion
The aim of the work presented in this chapter was to prove that microorganisms isolated from a
hemicelluloseenriched environment were able to utilize pentose sugars. In particular, the ability
to use pentose sugar carbon sources in the presence of glucose was considered to be a useful
property, due to the natural combination of these molecules in agricultural waste products,
which may serve as a fermentation feed-stock.
An additional focus of this chapter was to study and explore the metabolic activities of E-PUB.
This not only informs the interpretation of the results of the experiments reported in this
chapter, but also gives some additional information about the end-products of the different
metabolic processes that may be taking place, so informing future research.
Microbial metabolism consists of a network of biochemical processes to maintain life.
Metabolism consists of two major phases: catabolism and anabolism. Usually bacteria
breakdown sugars as a main carbon source or nutrient in the catabolism phase. Microorganisms,
usually heterotrophic bacteria, produce a range of by-products as a result of the anabolic phase.
The most common carbon source for bacteria is glucose, which is broken down via the major
carbohydrate-metabolizing pathways: Embden–Meyerhof–Parnas (EMP) pathway (glycolysis),
pentose phosphate (PP) pathway and the tricarboxylic acid cycle (TCA cycle).
Figure 4.3 shows the glycolytic pathway and its metabolic interconnection with the pentose
phosphate pathway and tricarboxylic acid cycle.

Chapter 4 Catabolic Characteristics of E-PUB
63
Figure 4.3: Metabolic pathways for the breakdown of pentose and glucose sugars (Ref: The new world encyclopedia (http://www.newworldencyclopedia.org/entry/Citric_acid_cycle) and Pelicano et al., 2006, page 4634, figure 1).
Legend: The solid arrows indicate glycolytic reactions and the dashed arrows show the pentose
phosphate pathway. The enzymes abbreviations are HK, hexokinase; PGI, phosphoglucose
isomerase; PFK, phosphofructokinase; TPI, triosephosphate isomerase; GAPDH, glyceraldehyde-
3-phosphate dehydrogenase; PGK, phosphoglycerate kinase; PGM, phosphoglycerate mutase;
PK, pyruvate kinase; PDH: pyruvate dehydrogenase; LDH: lactate dehydrogenase. The colour
schemes in the TCA cycle are as follows: enzymes, coenzymes, substrate names, metal ions,
inorganic molecules, inhibition, and stimulation. The enzymes required to breakdown xylose in
addition to glucose are glucose-6-phosphate dehydogenase, xylose reductase, xylitol
dehydrogenase.

Chapter 4 Catabolic Characteristics of E-PUB
64
Figure 4.4 shows a simplified scheme of the extra- and intracellular pathways involved in xylose
catabolism include enzymes from the glycolytic pathway.
Figure 4.4: Metabolic pathways for the breakdown of sugars (Xylose included)
Ref: Lu et al., 2010, page 3, figure 2.
Legend: An02g07470 (fructose-bis-phosphate aldolase); An14g04920 (triose phosphate
isomerase); An16g01830 (glyceraldehde-3-phosphate dehydrogenase); An08g02260
(phosphoglycerate kinase); An18g06250 (enolase); An07g09530 (pyruvate dehydrogenase
complex: pyruvate dehydrogenase); An07g06840 (dihydrolipoamide dehydrogenase); TCA cycle
enzymes: An08g10530 (aconitase); α-ketoglutarate dehydrogenas complex: An07g06840
(dihydrolipoamide dehydrogenase); An12g07850 (fumarase); An07g02160 (malate
dehydrogenase); enzymes from the pentose phosphate pathway: An02g02930 (ribose-5-
phosphate isomerase); An07g03850 (transaldolase); enzymes involved in anaerobic redox
balancing: An06g00990 (cytoplasmic fumarate reductase); enzymes involved in polyol
metabolism: An01g03480 (sorbitol dehydrogenase); acetate formation: An16g07110 (acetyl-CoA
hydrolase); xylose breakdown: An01g03740 (xylose reductase)
Xylose reductase, which is a pentose reductase and member of the aldoketoreductase family 2
(AKR2), catalyses the first step in five carbon metabolism by reducing xylose and arabinose to
xylitol and arabitol. The gene encoding xylose reductase (Texr) has been isolated from Candida
sp, Pichia stipitis and the thermophilic fungus Talaromyces emersonii.

Chapter 4 Catabolic Characteristics of E-PUB
65
Xylose catabolism involves a series of oxidation and reduction reactions to form D-xylulose,
which then enters the pentose phosphate pathway after phosphorylation to D-xylulose-5-
phosphate. The combined action of xylose reductase and xylitol dehydrogenase is required to
convert D-xylose to D-xylulose and all enzymes of the D-xylose pathway can be used in the L-
arabinose pathway, where arabitol is oxidized by NAD+-dependent arabitol dehydrogenase
producing L-xylulose. This is then converted to xylitol by NADPH-dependent L-xylulose reductase
(Jez and Penning, 2001).
Hahn-Hagerdal et al. (2001) improved the metabolic pathways essential to fermentative
capabilities of recombinant Saccharomyces strains, engineered for xylose metabolism. Several
xylose reductase genes have been identified from different sources (Amore et al., 1991; Billard
et al., 1995; Handumrongkul et al., 1998), and all show a common specificity for NADPH. Site-
directed mutagenesis and structural studies with Candida tenuis xylose reductase (Kratzer et al.
2006), a dual-specific enzyme with a preference for NADPH, revealed the main determinants
involved in pentose-specific substrate-binding recognition, key residues involved in coenzyme
interaction and suggested mechanisms by which certain AKRs can utilize both co-enzymes
(Petschacher et al.,2005; Di Luccio et al., 2006). Xylose reductase from Pichia stipitis, also
targeting amino acids involved in coenzyme interaction resulted in mutant proteins with
reversed coenzyme preference from NADPH to NADH in previous study (Watanabe et al., 2007).
Environmental pentose utilizing bacteria (E-PUB) bacteria were able to breakdown the sugars
tested, including a number of pentose sugars. Pentose sugars are broken down using the
pentose phosphate pathway and glucose is catabolised through the EMP pathway (Figure 4.3
and Figure 4.4). The metabolic pathway diagram illustrates the pathway that the E-PUB were
able to use, in order to utilize pentose naturally, due to presence of xylose reductase and xylitol
dehydrogenase enzymes, whereas previous research has described the need for genetically
engineering to activate of the gene encoding those enzymes. In the previous research, S.
cerevisiae XKS1 was used to initiate the evolution in continuous culture under aerobic conditions
with xylose and arabinose as limiting carbon sources (Karhumaa et al., 2006). Ethanol
productivity benefited from the design of stable S. cerevisiae strains that would co-consume
xylose and arabinose, since both sugars are present in lignocellulosic feedstock. So far there is
only one example of chromosomal integration of xylose and arabinose pathway genes in S.
cerevisiae (Jeppsson et al., 2003). In these engineered industrial strains (TMB3061 and
TMB3063) that carry the Pichia stipitis xylose pathway combined with the B. subtilis - E. coli
arabinose pathway the consumption of arabinose and xylose was low and ethanolic

Chapter 4 Catabolic Characteristics of E-PUB
66
fermentation was limited by extensive arabitol formation from XR enzyme (Karhumaa et al.,
2006). The E-PUBs are aerobic bacteria; however it was necessary to create an anoxic
environment in order to encourage them to produce amino acids.
E-PUB organisms were able to use pentose sugars as a carbon source during cultivation, without
any genetic modification. It has been previously reported that C. glutamicum had been
metabolically engineered to utilize the pentose sugar, xylose (Kawaguchi et al., 2006). The E-PUB
isolates were able to naturally breakdown xylose, arabinose and/or ribose as a sole carbon
sources.
The only isolate capable of utilizing all of the pentose sugars tested was Nocardia elegans (NC4-
1), which was isolated from the sugar treatment pond water of the Proserpine sugar mill, while
the other microorganisms used glucose and at least one pentose sugar. The utilization of both
hexoses and pentose sugars from lignocellulosic materials is promising for future sustainable
recycling, despite there being no previous publications regarding pentose usage from sugar
industry waste. Utilization of hexoses by bacteria and yeast is well known in fermentation
technology but the information of the ability of these organisms to utilized pentose sugars is
scant. The ability to ferment pentoses is not widespread among microorganisms and the most
promising yeast species identified so far are Candida shehatae, Pichia stipitis and Pachysolen
tannophilus (Hahn- Hagerdal et al., 2007; Chandel et al., 2010c). In order to improve the
suitability of these strains at an industrial level, many efforts have been made to construct
appropriate strains by cloning and expression of pentose metabolism genes into common hosts
such as Saccharomyces cerevisiae, Zymomonas mobilis, Escherichia coli etc (Jeffries, 2007; Hahn-
Hagerdal et al., 2007). However it is still remains a challenging issue to get suitable strains that
will fulfil the requirements of industrial production from lignocelluloses (Zhang et al., 2010).
Xylose was the most commonly used pentose sugar of those tested, while ribose supported the
growth of only one isolate. Arabinose, whilst being used by several isolates, did not permit
growth to the same extent, as measured by the final optical density, as other carbon sources in
most cases. The exception to this observation was Actinomyces odontolyticus (NC1-3), whose
growth with arabinose was virtually the same as with the other carbon sources tested.
The analysis of utilization of dual carbon sources was the second objective of this chapter. The
reason that biological treatments of industrial waste are so difficult is that the waste usually
contains many kinds of carbon sources. The efficient utilization of mixtures of various sugars is
critical for attaining complete conversion of lignocellulosic wastes. The physiological capability
for carbon catabolite repression mechanisms, present in many bacteria, is generally regarded to

Chapter 4 Catabolic Characteristics of E-PUB
67
be a mechanism that has evolved to ensure sequential carbohydrate utilization (Nampoothiri
and Pandey, 1995; Brückner and Titgemeye, 2002), with the most energy-efficient carbohydrate
being utilized first. While E. coli is able to ferment xylose, the utilization of this sugar by this
microbe during lignocellulosic hydrolysate fermentation is delayed and is often incomplete
(Naono et al., 1965; Dien et al., 2000). Likewise, genetically engineered Zymomonas mobilis has
been shown to preferentially utilize glucose during co-fermentation of sugar mixtures
(Mohagheghi et al., 2002).
Our research agrees with the findings of Singh and Mishra (1995). Since our E-PUB isolates were
able to utilize a combination carbon source consisting of glucose and a pentose sugar, the sugars
were used sequentially, with glucose used first followed by a lag period, and subsequently a
second exponential phase, thus demonstrating a typical catabolite repression. The bacteria used
in these experiments were not genetically modified to promote the preferential use of pentoses,
but are natural isolates. While that did not measure the levels of the remaining concentrations
of sugar substrates, both first and second growth curves were followed to a subsequent
stationary phase, which would normally suggest that the carbon source was depleted. It has is
predicted that all the sugars may be depleted after the two growth phases; however this study
only measured the glucose levels, which were very low, indicating that this sugar, at least, was
markedly depleted.
There is currently limited information in the literature about bacterial pentose-utilization. The
work presented in this chapter has proven that the isolated indigenous microorganisms were
able to metabolise pentose sugars. It was established that these organisms used pentose sugars
with approximately the same efficiency as glucose utilization when tested as single carbon
source.
However, as a part of a dual carbon source system, the pentose sugars in some cases appeared
to be less efficient as a source of carbon than they were in a single carbon source test. This may
be partially due to a technical anomaly, attributed to the difficulty in accurately measuring the
optical density of slow growing organisms. However, the fact that two of the species were able
to use xylose more effectively as a dual-carbon source than as a single one, confirms our
confidence in these species as novel biofermentation starter cultures.
As previously discussed, pentose-utilization is an important, but rare, trait for the economically
feasible production of chemicals from lignocellulosic biomass by microbial cells. This limitation
was resolved in this study by investigation of indigenous organisms, which proved to be naturally
capable of efficiently and concurrently metabolizing both glucose and xylose.

Chapter 4 Catabolic Characteristics of E-PUB
68
The question may be raised as to the identity of the limiting factor, which causes the diauxie
effect to occur. This factor may be: carbon source; nitrogen source or phosphorous, or indeed
growth conditions such as oxygen supply or pH. This have shown that the glucose levels drop to
a very low figure (Table 4.5) and so predict that carbon source is indeed the limiting factor. Were
the nitrogen or phosphorus to be limiting factors, a second growth phase would not be
observed, since there were no alternative supplies of these elements to fuel such a phase. The
only nutrient supplied in two forms, was the carbon source, and so this is the most likely factor
to be responsible for the diauxie effect.
4.5 Conclusions
Environmental pentose-utilizing bacteria (E-PUB) were able to metabolise pentose sugars when
they were presented as a single carbon sole in minimal growth media, under standard aerobic
conditions. In addition the E-PUB used a diauxie growth pattern, typically observed as a result of
catabolite repression, when presented with a dual sugar carbon source consisting of glucose and
a pentose sugar. In most cases they continued on to utilize the pentose sugar present, after the
glucose supply had been exhausted. As Nocardia elegans also has the ability to use any of the
pentose sugars tested, it is a very promising candidate for industrial fermentations.

Chapter 5 Anabolic Characteristics of E-PUB
69
CHAPTER 5
Anabolic Characteristics of Environmental Pentose-
Utilizing Bacteria
5.0 Summary
In order to determine the ability of environmental pentose-utilizing bacteria (E-PUB) to produce
commercially valuable products such as amino acids from a process of fermentation of pentose
sugars, E-PUB isolates were grown in various pentose substrates, and the presence of amino
acids was analysed using an HPLC system. The products varied, with one or two amino acids
being recovered in all cases.
When the E-PUB isolates were tested in a diauxie system, with dual sugar substrates consisting
of glucose (hexose) plus one of the pentose sugars, the end-product results showed a single
predominant amino acid, identified as glycine by comparison with the standards. This amino acid
is widely used for medical and industrial purposes, and is considered to be a very useful by-
product of the fermentation process, using E-PUB with dual sugars (glucose plus pentose) as
substrates.
5.1 Introduction
Microorganisms closely related to the Corynebacterium genus are frequently used for
fermentation purposes because they are able to utilize various sugars as substrates and in
addition, Corynebacterium strains can produce valuable amino acids as end-products. There
have been several studies performed regarding amino acid production from pentoses, using
genetically modified Corynebacterium strains (Ikeda and Katsumata, 1999; Moritz et al., 2000;
Ohnishi et al., 2005).
Biotechnological production processes have been used for the industrial production of amino
acids for the last 50 years. Market development has been particularly dynamic for the flavour-
enhancer, glutamate and the animal feed amino acids, L-lysine and L-threonine, which are
produced by Corynebacterium spp from hexose sugar sources such as sucrose or glucose. The
market for synthetic amino acids is becoming increasingly important, with annual growth rates
of 5-7% (Leuchtenberger et al., 2005). Amino acids are useful as building blocks for active
ingredients in the production of pharmaceuticals, cosmetics and agricultural products. Nutrition
and health will continue to be the driving forces for exploiting the potential of microorganisms,

Chapter 5 Anabolic Characteristics of E-PUB
70
and possibly also of suitable plants, in order to arrive at even more efficient processes for amino
acid production.
Wise selection of raw materials is essential for economic amino acid production, especially since
the carbon source represents a major component of variable production costs. This explains the
association between amino acid and sugar manufacturing which has developed over time. Some
amino acid production plants are located geographically close to sugar mills in order to decrease
transport costs and on occasion, joint ventures are formed. Depending on the geographical
location of the plants, carbon sources such as cane molasses, beet molasses, or starch
hydrolysates from corn, potato or cassava are used. While molasses is common in Europe, South
America and China, starch hydrolysate is the most important carbon source in North America.
Tapioca hydrolysate, the starch hydrolysate from cassava is widespread in South-East Asia.
However, pure sugars are favourably compared with molasses because of the unwanted side
reactions and variable qualities of the complex media components.
Earlier studies (John et al., 2007) concluded that sugar derivatives from the cellulose and
hemicellulose components of plant biomass are an attractive option as sustainable substrates
for the biological production of many organic acids, fuel and other industrial chemicals. With
respect to the pentose-fraction of hemicellulose biomass, there are several microorganisms
available that have been genetically engineered to break down pentose sugars to produce
ethanol and other organic chemicals. However, this approach is limited by the cost of
production, the physiological characteristics of the microorganisms and the lack of public
support for the use of genetically engineered microbes in large scale chemical production
(Ronald and Admachak, 2008).
This negative public attitude could be addressed by using natural isolates, screened for their
capacity to break down pentose sugars. The alternative process, of selecting mutants that
switch off the glucose metabolism so that alternative, available substrates such as pentose
sugars would be used instead, retains some difficulty in terms of public negativity, as the
mutation aspect is not currently in favour (Ronald and Admachak, 2008).
The main focus of this chapter is to ascertain whether naturally occurring Corynebacterium spp,
now shown to be able to utilize pentose sugars, are also able to produce any industrially
valuable amino acids.
The aim of this current study was to isolate, test and propagate microbes that could effectively
use pentose sugars naturally, and subsequently produce economically valuable end-products. As
the target end-product for this research was to be an amino acid and Corynebacterium spp are

Chapter 5 Anabolic Characteristics of E-PUB
71
known to be able to produce amino acids, this group, and environmental organisms
phenotypically similar to it (see Chapter 3), that are known to utilize pentose sugars (see Chapter
4), were further investigated to identify their end-products, in this chapter.
5.2 Materials and Methods
5.2.1 Equipment
The analysis was performed using two identical Agilent 1100 HPLC (Heracles, Japan) systems.
Each system consisted of a binary pump, a UV detector, a fluorescence detector and an auto
sampler. A reverse phase Agilent Zorbax Eclipse C18 column AAA (4.6150 mm, 3.5 micron) was
used for the chromatographic separation.
5.2.2 Chemicals
Amino acid standard solutions were obtained from Agilent (P/N 5061-3330). O-phthalaldehyde
3-mercaptopropionic acid (OPA-3MPA) and borate buffer were from Agilent. OPA-3MPA was
stored at 2–8°C in small vials crimped with silicon rubber, PTFE-coated cap (Woodward, 2007;
Henderson, 2009). A fresh aliquot of OPA was used for each set of samples. All other solvents
were HPLC grade from Agilent. The HPLC separation of the derivatized amino acids required two
mobile phases. The mobile phases, A (40 mmol/l Na2HPO4 at pH 7.8), and B (45% acetonitrile,
45% methanol, 10% water) were filtered through a 0.22-micron Millipore Durapore PVDF
membrane filter. To prevent microbial growth in the mobile phase, sodium azide (5 mg/L) was
added (Woodward et al., 2010). Potassium tetraborate, perchloric acid and potassium
dihydrogen phosphate were used for protein precipitation and extraction in this experiment.
5.2.3 Cultivation and sample preparation
As all of the E-PUB organisms were able to break down xylose as a carbon source in the growth
medium, xylose was the prefered substrate to analyse the end products. . Duplicate microbial
isolates were cultured on LB media with xylose as a carbon source for 3 days in shaking-
incubator. These young cultures were then held for 24 hours unshaken to produce sufficiently
anoxic conditions to encourage the production of the required end-products. Aliquots (50 μL) of
cultures were centrifuged to separate pellets at 5000 rpm (4620×g) for 30 min. According to the
method developed by Frank and Powers (2007), samples were added to an equal volume (20 μL)
of a standard (62.5 μmol/L) and HPLC-grade water (160 μL) for a final volume of 200 μL. Proteins
were subsequently precipitated by adding 200 μL of 0.5 mol/L perchloric acid. After protein
precipitation, the samples were centrifuged at 15,000 × g for 5 min at room temperature. 150 μL
of the supernatant was filtered in a Spin-X 0.2-μm micro-centrifuge filter by centrifugation at

Chapter 5 Anabolic Characteristics of E-PUB
72
15,000 × g for 1 min. 100 μL of the filtered sample was collected and split between two sample
vials to minimize the time between derivatization and injection on to the HPLC system.
The microbial extraction methods for amino acid analysis require a wide range of solvents. Some
examples include the use of ethanol perchloric acid, glycerol and an aqueous solution of
trichloracetic acid (Nunn and Keil, 2006). The perchloric acid extraction method is a considerably
more effective for qualitative analysis of amino acids than all other extraction techniques. In
addition, the use of perchloric acid is not expensive compared to other extraction chemicals.
The weight of the dry biomass was measured using a 100 mL sample, which was centrifuged in
100 mL centrifuge tubes (4°C, 25 minutes at 5000 rpm(4620×g)), and washed three times with
30 mL distilled water. The resuspended washed pellet was placed in an aluminum jar and freeze
dried at -50°C, vacuum pressure 300 p.a. The weight of the dried pellet was measured to permit
calculation of the yields per unit weight of glucose.
5.2.4 Standard preparation
All amino acid standards were stored at −20 °C. The linearity of the response for each individual
standard across different concentrations ranging from 5 to 1000 μmol/L was measured by
plotting the peak area for each amino acid divided by the area of the internal standard vs.
concentration (Frank and Powers, 2007).
5.2.5 Chromatographic system
The UV detection was performed with an absorbance wavelength of DAD (Dicode Array
Detector) 215 nm. The flow rate of the mobile phase was 1 ml/min throughout the analysis. The
total HPLC run time for the separation of the derivatized amino acids in a single sample or
standard was approx 6-8 min per sample.
Automatic pre-column derivatization with OPA-3MPA was performed at room temperature,
according to the injector programmes listed in Table 5.1, using 1 µL of filtrate (Woodward,
2007). After the derivatization, 0.5 µL of the mixture were injected for each chromatographic
separation. Primary amino acids were derivatized with OPA-3MPA and detected by a UV
detector at 215 nm, with a reference=360 nm, band width=10, slit of 4 nm, peak width of N 0.1
min. The protocol of HPLC injector programme is detailed in Table 5.1.

Chapter 5 Anabolic Characteristics of E-PUB
73
Table 5.1: HPLC injector programme for amino acid detection
5.3.6 Qualitative and quantitative analysis
Performance of a quantitative analysis requires the preparation of a calibration curve (see
Appendix Figure B1 a-e). The quantification was calculated as the average peak area per µmol of
amino acid present in the standard. To undertake quantitative analysis of amino acids, a sample
of a known volume is injected, and the peak area was calculated. The height and area of a peak
are proportional to the concentration of the corresponding component. A calibration curve (see
Appendix Figure B1 a-e) was created using the standard sample.
5.2.7 Recovery and variability
The recovery of the amino acids was calculated as the difference between spiked and unspiked
samples. Each sample was tested in triplicate and calculated for their average recovery.
Recovery was expressed as [(found concentration−basic concentration)/spiked
concentration]×100%. The intra-assay coefficient of variation (CV) was determined by replicate
analysis of a standard solution (n = 10) and a quality control sample (n = 10) in a single run (Frank
and Powers, 2007).
The between-run CV was 10% for all amino acids studied. A critical step for the reproducibility of
the method was the addition of column washes, performed before every injection, with a
Injector Program for mobile phase A and B
Line Function Amount Reagent
1 Draw 2.5 µL Borate Buffer(pH=7.8)
2 Draw 0.5 µL Sample/standard
3 Mix 3 µL in washpot, 400 µL/min speed, 6
times
4 Wait 0.2 min
5 Draw 0.5 µL OPA-3MPA
6 Mix 3.5 µL in washpot, 400 µL/min speed, 6
times
7 Draw 32 µL Water as an injection diluent
8 Mix 3 µL in washpot, 400 µL/min speed, 8
times
9 Inject 0.5 µL

Chapter 5 Anabolic Characteristics of E-PUB
74
mixture of methanol/water (90/10) to regenerate the column. The method demonstrated good
chromatographic separation of amino acids.
5.3 Results
5.3.1 Amino acid standard analysis
There was a linear relationship between the area and the amount of sample in the standard
curve. Figure 5.1 shows the separation of amino acid standards with mobile phase A at pH 7.8.
Separation and detection of a standard mixture (Agilent) of aspartic acid, glutamic acid,
asparagine, histidine, glycine, threonine, arginine, alanine, cysteine, tryptophan, phenylalanine,
and lysine were analysed with this mobile phase. The analysis of amino acids in physiological
fluids has been undertaken with the use of OPA derivatives. The OPA derivatized samples are
detectable in the UV range. The elution times are noted in Table 5.2.
Figure 5.1: Chromatogram of a mixture of 0.5 µL amino acids standards separated using HPLC
with mobile phase A (pH=7.8).
The range (1-13) of detectable standard amino acids is shown in Table 5.2
Table 5.2: Names of amino acids detected and their elution times
Peak no Name of amino acid Elution
time
(min)
Peak no Name of amino acid Elution
time
(min)
1 Aspartic acid 0.2 8 Alanine 3.3
2 Glutamic acid 0.5 9 Tyrosine 3.8
3 Serine 2.1 10 Cysteine 4.0
4 Histidine 2.6 11 Phenylalanine 4.4
5 Glycine 2.7 12 Isoleucine 4.6
6 Threonine 2.8 13 Lysine 4.8
7 Arginine 3.1
5.3.2 Amino acids detected from E-PUB metabolic process using pentose sugar
substrates
1 2 3 4 5 6 7 8 9 10 11 12 13

Chapter 5 Anabolic Characteristics of E-PUB
75
The microbial cells were treated as described above, and subjected to amino acid analysis (also
as described above). All amino acids were separated and identified with both mobile phases and
the within-run and between-run imprecision were comparable in both samples. Amino acids
were separated and identified using chromagraphic separation method with mobile phase A.
The separating condition was a combination of mobile phase of 45% acetonitrile, 45% methanol,
10% water in a flow rate of 1.0 ml/min. These amino acids were separated well under these
conditions. There were few obvious peaks of other components in the sample could be seen in
the chromatogram.
Figure 5.3 shows that all of the tested cultures: N. elegans (NC4-1); A. odontolyticus (PC4-1, NC1-
3); C. glutamicum (NC1-2, PC1-2; and P. freudenreichii (31TG) produced amino acids from xylose
metabolism and also after diaux. A variety of amino acids was produced in quantifiable amounts
after pentose sugar utilization.
Table 5.3: Amino acids produced by E-PUB isolates
Isolate Amino acid from single sugar pentose substrate
Concentration mg/L
N. elegans (NC4-1) Threonine 36
A. odontolyticus (PC4-1, NC1-3) Arginine 45
Cysteine 6
C. glutamicum (NC1-2, PC1-2)) Arginine 46
Cysteine 3
Glycine 5
P. freudenreichii (31TG) Arginine 47
Cysteine 10
Glycine 5
Alanine 6

Chapter 5 Anabolic Characteristics of E-PUB
76
a. N. elegans (NC4-1)
b. A. odontolyticus (PC4-1)
c. C. glutamicum (NC1-2)
d. P. freudenreichii (31TG)
e. A. odontolyticus (NC1-3)
Figure 5.2: Continued on next page

Chapter 5 Anabolic Characteristics of E-PUB
77
f. C. glutamicum (pc1-2)
Figure 5.2: Chromatograms of the amino acid mixture present in microbial cultures. Samples derived from the set of E-PUB cultures, and separated using HPLC with mobile phase A (pH=7.8) (A) photodiode array UV detection at 215 nm.
As suggested by Frank and Powers (2007), the within-run CVs for all detectable amino acids
studied was <7% (Table 5.4). The between-run coefficient of variation (CV) was <10% for all
amino acids studied. A critical step for the reproducibility of the method was the addition of
column washes. These were performed by single injections, with a mixture of 45% acetonitrile,
45% methanol and 10% water to regenerate the column.
The method uses minimal sample volume and derivitization of amino acids with o-
phthalaldehyde and UV detection. Recovery and variability data are shown in Table 5.4. For all
concentrations, recovery of amino acids was between 94 and 96%, intra-assay coefficient of
variation (CV) was 1–6%, and inter-assay CV was 3–11% (Table 5.4). The results indicated a good
recovery of all amino acids which was acceptable.
Table 5.4: Performance characteristics of HPLC UV detection of amino acid
Between run and within run coefficient of variation (CV) for amino acids
Amino acids Elution time (min)
Within-run, %CV
Between-run, %CV
Recovery %
Arginine 2.1 5.9 10 96
Glycine 2.6 6.5 10.03 95
Threonine 2.7 6.3 10 94
Alanine 3.4 2.5 4.8 94
Cysteine 4.0 2.2 4.9 94

Chapter 5 Anabolic Characteristics of E-PUB
78
The wide analytical measurement range for all the amino acids studied could only be achieved
using UV detection, with the exception of hydroxyproline and proline. Since these amino acids
were detected by derivatizing with FMOC and using a fluorescence detector, a lower linearity
range was obtained. The within-run coefficient of variation (CV) for all amino acids studied was
5% (Table 5.2).
Figure 5.3 shows the analysis of an LB medium blank. It demonstrated that no free amino acids
were present in the medium. Those peaks that were present were all near 0 or below 1 mAU
value, thus being of no significance in terms of the final analysis.
Figure 5.3: HPLC for LB medium-blank analysis
The LB medium contains yeast extract, which is an inexpensive organic source of proteins and
vitamins for cell growth and synthesis of enzymes such as amylase and protease. This had the
potential to interfere with the results; however the HPLC of the blank medium showed that free
amino acids were not present in the medium. It may be that the yeast extract contains peptides
and small molecular weight protein components, but not significant amounts of free amino
acids.
5.3.3 End product analysis of diauxie metabolism
The end-products after dual-substrate cultivation, using glucose plus a pentose sugar (xylose,
arabinose and ribose) as substrates, were analysed using the HPLC methods as noted above.
Figure 5.4 a shows that four isolates, A. odontolyticus (PC4-1 andNC1-3), N. elegans (NC4-1) and
P. freudenreichii (31TG), cultured on a glucose and arabinose combination, produced the same
peaks over the 7 minutes of HPLC runtime. Glycine was separated from the extracted products
of growth of A. odontolyticus (PC4-1 and NC1-3), N. elegans (NC4-1), and P. freudenreichii (31TG)
isolates. There was a sharp unknown peak at 1.3 min (Figure 5.4 b). However, as shown in
Figure 5.4 c, all six indigenous microorganisms N. elegans (NC4-1), A. odontolyticus (PC4-1 and

Chapter 5 Anabolic Characteristics of E-PUB
79
C1-3), C. glutamicum (PC1-2 and NC1-2), P. freudenreichii (31TG) produced glycine as their
major end product after growth on the glucose with xylose as a dual sugar combination medium.
5.4a
5.4b
5.4c
Figure 5.4: End-products of dual-sugar metabolism (glucose and pentose) 5.4a: End product analysis as produced using glucose plus arabinose substrate combination, by isolates A.odontolyticus (PC4-1), A.odontolyticus (NC1-3), and N.elegans (NC4-1), and P. freudenreichii (31TG). 5.4b: End product analysis as produced using glucose plus ribose substrate combination, by isolate N. elegans (NC4-1). 5.4c: End product analysis as produced using glucose plus xylose substrate combination, by isolates C. glutamicum (PC1-2), C. glutamicum (NC1-2), A. odontolyticus (NC1-3), A. odontolyticus (PC4-1) and N. elegans (NC4-1).
All E-PUB isolates produced approximately 22 mg/L glycine after dual sugar xylose+ glucose
utilization. N. elegans (NC4-1) and A. odontolyticus (PC4-1, NC1-3) produced 22 mg/L glycine
after arabinose + glucose utilization as a combination carbon source. P. freudenreichii (31TG)

Chapter 5 Anabolic Characteristics of E-PUB
80
also produce the same amount of glycine after ribose + glucose utilization. These results are
shown in Table 5.5, together with the results of xylose metabolism for comparison.
Table 5.5: Amino acid production using single and dual carbon sources as substrates
Isolate Amino acid from single sugar substrate
Concentration mg/L
Amino acid from dual sugar substrate
Concentration mg/L
N. elegans (NC4-1) Threonine 36 Glycine 22
A. odontolyticus (PC4-1, NC1-3)
Arginine 45 Glycine 22
Cysteine 6
C. glutamicum (NC1-2, PC1-2))
Arginine 46
Cysteine 3
Glycine 5 Glycine 22
P. freudenreichii (31TG) Arginine
47
Cysteine 10
Glycine 5 Glycine 22
Alanine 6
The measurements of dry biomass after single and dual substrates utilized are given in Table 5.6.
Total biomass was measured following utilization of a single carbon source, and also following
the use of a combination of hexose and pentose (glucose and xylose, glucose and arabinose,
glucose and ribose) substrates during bacterial cultivation.
Table 5.6: Total biomass measured from single pentose and dual sugar carbon substrate
While the amounts of amino acid produced appeared to be similar, it was only by calculating the
yield of amino acid with respect to the biomass production in each case, that it became possible
Biomass Yx/s (g/g) (Single Pentose sugar carbon source)
Biomass Yx/s (g/g) (Dual sugar carbon source)
Bacteria Xylose Arabinose Ribose Glucose/
Xylose
Glucose/
Arabinose
Glucose/
Ribose
C. glutamicum (PC1-2) 0.48 - - 0.44 - -
C. glutamicum (NC1-2) 0.45 - -
0.51 - -
A. odontolyticus (PC4-1) 0.42 0.46 -
0.37 0.41 -
A. odontolyticus (NC1-3) 0.50 0.51 -
0.51 0.48 -
N. elegans (NC4-1) 0.54 0.54 0.51
0.55 0.52 0.49
P. freudenreichii (31TG) - 0.52 -
- 0.55 -

Chapter 5 Anabolic Characteristics of E-PUB
81
to determine which cultures were more efficient in an industrial context. The various yields of
amino acids, expressed in mg amino acid per gram of biomass (calculated on the basis of 500mL
of culture) are summarized in Table 5.7 below.
Table 5.7: Yield of amino acid expressed in mg per g of biomass, based on a 500mL culture size
5.4 Discussion
By means of this investigation, successful anabolism was detected after use of pentose sugars by
isolated indigenous microorganisms.
Among various cheap carbon sources, industrial by-products such as molasses and other
cheaper components like wheat bran and sugar cane bagasse have industrial potential, since
they support both biomass increase, and enzyme production (Kanekar et al., 2002). Sugar cane
bagasse has been the particular focus of this work.
Microorganisms are known to convert nutrients to various vital components necessary to their
survival and reproduction. With the fermentation method, raw materials are added to
microorganism culture media, and the proliferating microorganisms are encouraged to produce
E-PUB Amino acid Amino acid mg/500 mL (Single sugar)
Amino acid Yield (mg/g)
Amino acid mg/500 mL (Dual sugar)
Amino acid Yield (mg/g)
N. elegans (NC4-1) Threonine 18 33.3
Glycine 11 20
A. odontolyticus (PC4-1) Arginine 22.5 48.9
Cysteine 3 6.5
Glycine 11 29.7
A. odontolyticus (NC1-3) Arginine 22.5 44.1
Cysteine 3 5.8
Glycine 11 21.5
C. glutamicum (NC1-2) Arginine 23 47.9
Cysteine 1.5 3.1
Glycine 2.5 5.2 11 25
C. glutamicum (PC1-2) Arginine 23 51.1
Cysteine 1.5 3.3
Glycine 2.5 5.5 11 21.5
P. freudenreichii (31TG) Arginine 23.5 45.1
Cysteine 5 9.6
Glycine 2.5 4.8 11 20
Alanine 3 5.7

Chapter 5 Anabolic Characteristics of E-PUB
82
amino acids by manipulation of the physical conditions. The enzymes play an important role in
catalysing chemical reactions, are indispensable to degrade and synthesize substances and
various amino acids are produced as a result of these reactions.
Figure 5.5: Formation of intracellular and extracellular amino acid (Hundal and Taylor, 2009, page E604, figure 1)
In previous, related research, McCowan and Phibbs (1974) found that the total intracellular pool
increased during exponential growth of Bacillus licheniformis and then decreased rapidly after
the end of growth. Of great interest to the present research, they found that most of the amino
acids were present at low concentrations, while glutamate and alanine comprised 60 to 90% of
the total intracellular free amino acid at most times during the growth cycle. It was concluded
that, in addition to providing monomers for protein synthesis, the intracellular amino acid pool
could be maintained for the storage of energy-providing metabolic intermediates (Hundal and
Taylor, 2009). A 10-fold increase in extracellular amino acid was observed as the cells changed
from vegetative to sporulation metabolism, mostly due to the extrusion of intracellular amino
acid. The bacterial isolates used in the current research were not capable of sporulation,
however it is considered most likely that the maximum production of amino acids was also
undertaken in the stationary phase of their growth cycle, and in the anaerobic conditions.
The main focus of this research was to study amino acid production by E-PUB, with intracellular
amino acids as the primary target, these being indicative of internal metabolic processes related
to the synthesis of the bacterial cell wall. Microorganisms produce a variety of amino acid
products in order to build the cell wall, following which excess amino acids are excreted as

Chapter 5 Anabolic Characteristics of E-PUB
83
extracellular amino acids (Figure 5.5), (Scheffers and Pinho 2005). In this research, it was decided
to focus on intracellular amino acids, as per the method published by Frank and Powers (2007).
So intracellular amino acid extraction was performed from the centrifuged pellet, rather than
from the supernatant. While it is possible that the measured levels could have been higher, if
the supernatant had also been tested, this was not done, as it was expected that a greater
concentration of amino acids would be expected intracellularly, this being their primary
destination.
Ideally, further optimization of various growth parameters such as pH, temperature, incubation
time, levels of nitrogen, phosphorous and other growth factors and the oxygen levels, would
have been performed, preparatory for upscaling the process to an industrial level. Such
optimization would no doubt have the result of improving the yield, which was rather low in the
reported results. It is also acknowledged that further extraction and analysis of extracellular
amino acids, potentially present in the culture supernatant, may prove to be a factor in
improving the yield.
These trials produced good yields of amino acids compared to previous research. For example,
3.6 g/liter threonine was produced from the modified whey permeate by using Escherichia coli
ATCC 21151 for threonine production, and utilizing lactose as a carbon source (Young and
Chipley, 1983). Arginine concentration of the batch fermentation was 36.6 g/liter produced by
Corynebacterium crenatum (Xu et al., 2009). According to Dhillon et al., (1987) Bacillus
sphaericus produced 7g/ litre L-cysteine.
To determine the relationships between the processes of uptake, intracellular pool formation,
and incorporation of amino acids into protein, research regarding the quantitative and
qualitative analysis of bacterial intracellular free amino acids was examined. Significantly, not all
the amino acids present in the proteins of bacteria were present in the pools in detectable
concentrations (that is, greater than 0-1 mM). In particular, the pools of both Gram-positive and
Gram-negative bacteria were found to generally lack tryptophan, tyrosine, phenylalanine,
histidine, arginine, cysteine and methionine. Of the amino acids regularly present in bacterial
pools, leucine, isoleucine, serine, glycine, aspartate, valine, lysine and proline were frequently
present in low concentrations. Whereas according a previous report, 3.4 mM Glycine was
produced by Bacillus subtilis, 1.5 mM B. megaterium and 7.1 mM B. polymyxa; 3.1 mM alanine
produced by Bacillus subtilis, 0.6 mM B. megaterium and 20 mM B. polymyxa; 2.6 mM threonine
from Bacillus subtilis, 0.5 mM B. megaterium and 21 mM B. polymyxa (Tempest and Meers 1970;
Kirchman and Hodson 1986; Iwatani et al., 2007).

Chapter 5 Anabolic Characteristics of E-PUB
84
In the present study, following diauxie breakdown of a pentose sugar and glucose, the only
amino acid produced in measureable quantity was glycine. A. odontolyticus (PC4-1) yielded 29.7
mg/g of biomass, while NC1-3 produced 21.5mg/g biomass. Similarly, C. glutamicum (PC1-2)
produced 21.5mg/g biomass while NC1-2 produced 25mg/g biomass. P. freudenreichii and
N.elegans both produced the lowest concentrations of glycine, which was 20mg/g biomass. The
presence of glycine is not surprising, as it has been shown above to be a major end-product of
bacterial metabolic processes, and it is an end-product of amino acid productive pathways.
The amino acid spectrum produced after single pentose sugar growth was broader, with other
amino acids being detected, such as threonine, cysteine alanine and arginine. Of these, the best
yield was found with arginine, with figures of up to 51.1mg/g biomass produced by C.
glutamicum (PC1-2). However, since the context of this study was that of a mixed substrate, it is
unlikely that the production of arginine would be amenable to industrialization.
Variations in growth conditions were, whilst not explored fully in the present work, admitted to
be very important in studies of this kind. Obviously, the macromolecular composition and
metabolic activity of microorganisms may vary with changes in their growth environment
(Herbert, 1961; Neidhardt, 1963; Brown & Rose, 1969; Tempest, 1970). These phenotypic
changes in cell structure and functioning reflect changes in genetic expression and are
presumably mediated by some environmentally linked mechanism. Thus, changes in the growth
conditions must affect primarily the intracellular concentrations of substances reacting directly
with the genetic control mechanisms. In particular, intracellular amino acids are extremely
variable and markedly dependent on the nutritional complexity of the growth medium (Herbert,
1961; Neidhardt, 1963; Brown & Rose, 1969; Tempest, 1970).
In this work, E-PUB microorganisms have been studied for use in the production of various
industrially important amino acids. One very important variable in this type of work is that of
oxygen availability. Under conditions of oxygen deprivation, microbial cell growth (biomass
increase) is arrested, but the cells retain the capability to produce end-products including
organic acids, such as lactic acid, succinic acid, acetic acid (Inui et al., 2004). This allows, under
anoxic conditions, following aerobic culture, a bioprocess resulting in the production of
commercially valuable amino acids. Actinomyces and Propionibacterum are microorganisms
known to produce succinic and lactic acids (Siqueria, 2003) and so are good candidates for
industrial processes.
Further, oxygen requirements for the growth and production of end-products are known to be
influenced by osmotic pressure. Several researchers (Varela et al., 2002; Morbach and Krämer,

Chapter 5 Anabolic Characteristics of E-PUB
85
2003) have studied the influence of osmotic pressure on the growth and physiology of C.
glutamicum and related species. C. glutamicum was studied as it was considered a very
important microorganism for industrial level amino acid synthesis. Specific growth rates and
carbon-to-biomass yields decreased nearly linearly with increasing osmotic pressure, while the
rate of sugar uptake increased significantly. These scenarios may happen with E-PUB during their
metabolism. Neither of these studies was conducted with pentose sugar substrates, so that it
was predicted that, the response of the E-PUB on metabolic network to osmotic pressure may
be only assumed by comparison with the glucose substrate that was used.
Nitrogen is required for the growth of most bacteria, including Corynebacterium spp, Nocardia
spp and Actinomyces spp. Studies by Varela et al. (2002) demonstrated the nutritional
requirement for nitrogen by C. pyogenes, a bacterium that is quite different to other
Corynebacterium spp of animal origin. Cell wall composition, biochemical properties, metabolic
products and the absence of mycolic acids are the main differences from other species.
However, according to the vitamin requirement, this bacterium is very similar to Actinomyces
spp. Thus it is considered probable that the E-PUB will have a nitrogen dependant metabolism,
un-noted in the present research as nitrogen was provided in excess of demand.
In the 1950s, C. glutamicum was found to be a very efficient producer of L-glutamic acid. Since
that time, biotechnological processes with bacteria of the species Corynebacterium have
become very important in terms of tonnage and economic value. L-glutamic acid and l-lysine are
bulk products and l-valine, l-isoleucine, l-threonine, l-aspartic acid and l-alanine are among other
amino acids produced by Corynebacterium (Hermann, 2003). To date, l-cysteine has been
produced almost completely by extraction from biological materials such as hair. However, an
alternative bioprocess was established in 2000 using E. coli (Daßler et al., 2000). Several
coryneform strains were described that would produce significant amounts of l-cysteine (Wada
et al., 2002). The present research has shown that C. glutamicum produced arginine, glycine and
cysteine from the pentose sugars. The production of arginine and glycine from C. glutamicum
without genetic modification is a new finding. It is considered that the growth of this organism
on pentose sugars is what makes the observed difference in outcome. This will be the milestone
towards industrial production of amino acid in terms of saving costs.
This research did not continue to the extent of examining the enzymes present in E-PUB to
manufacture amino acids; however it is of interest to note the extent of current knowledge in
this field, as this may be quite relevant to industrial scale production. Amino acids can be divided
into groups on the basis of end-product production by anabolic pathway. Alanine is a part of the
pyruvate family whereas glycine and cysteine are members of the serine family; threonine is a

Chapter 5 Anabolic Characteristics of E-PUB
86
part of the aspartate family and arginine falls under the glutamate family. In the anabolic phase,
arginine and threonine are producd as end products through the TCA cycle.
Glycine, the major end-product of the E-PUB metabolism, is formed from 3- phosphoglycerate
(Figure 5.6), and arginine is produced from α-ketoglutarate. All of the environmental pentose-
utilizing bacteria (E-PUB) produced arginine as an end product, with the exception of N. elegans
(NC4-1), which produced threonine. This may be due to the presence of threonine synthase
enzyme. A. odontolyticus (PC4-1 and NC1-3), C. glutamicum (PC1-2 and NC1-2), and P.
freudenreichii (31TG) produced arginine, indicating the probable activity of a combination of the
enzymes arginino-succinate synthetase and arginino-succinate lyase (Figure 5.6). These bacteria
also have produced cysteine because of the possible presence of cystathionine lyase. C.
glutamicum (PC1-2 and NC1-2- ) and P. freudenreichii (31TG) produced glycine due to the
probable presence of glycine hydroxymethyltransferase enzyme. Only P. freudenreichii (31TG)
excreted alanine, this may be due to the presence of alanine transaminase.
In the process of amino acid production, there is an interim step, which is the formation of
glutamate. Although glutamate was not identified by HPLC as an end product, probably due to
the low concentrationm it was found in the review of published literature (chapter 2) that
Corynebacterium sp is mostly known as a glutamic acid producer organism. However in this
study it appeared that glutamate was not present and predictable cause may be that it was
further metabolised to arginine.
Figure 5.6: Glycine biosynthesis (http://themedicalbiochemistrypage.org/amino-acid-
metabolism.php#arginine)
According to Hui et al, (2001) and Thiele et al., (2002) applications of amino acids either in food
and pharmaceuticals or in animal feed nutrition, are expected to increase in the next few years.
It was predicted that, all 6 E-PUB isolates produced arginine and all except N. elegans (NC4-1)

Chapter 5 Anabolic Characteristics of E-PUB
87
produced cysteine. According to previous research (Hui et al., 2001), cysteine was mainly the l-
enantiomer, which is a precursor in the food, pharmaceutical and personal care industries. One
of the largest applications is the production of flavouring products, and arginine is also used as a
food supplement.
In the current research, C. glutamicum (NC1-2 , PC1-2) and P. freudenreichii (31TG) produced
glycine when a single pentose carbon source was metabolised. Glycine is very important in
pharmaceutical applications (Kuan et al., 2003). Furthermore, glycine was the major end product
from diauxie metabolism of all indigenous isolates. Metabolism of glycine may influence the
availability of substrates for the production of other amino acids. Glycine is one of glucogenic
amino acids, which refers to their ability to provide glucose to the blood. Additionally, glycine
also plays a role as a major inhibitory neurotransmitter in the spinal cord and brain stem, and an
anti-inflammatory, cytoprotective, and immune modulating substance (Gundersen et al., 2005).
The production of glycine by E-PUB would be a significant contribution to industrial amino acid
production in terms of health products.
5.5 Conclusions
After catabolic reaction with the pentose sugar, xylose, as a substrate, the isolate N. elegans
(NC4-1) produced threonine whereas A. odontolyticus (PC4-1 and NC1-3), C. glutamicum (PC1-
2and NC1-2), produced arginine and cysteine. Isolate P. freudenreichii (31TG) produced glycine,
arginine, alanine and cysteine after utilization of arabinose as a sole carbon source. All six
isolates produced glycine after dual (xylose and glucose) sugar catabolism. P. freudenreichii
(31TG) also produced glycine after metabolizing a dual sugar combination of glucose and
arabinose. The production of glycine is important because of its pharmaceutical significance.

Chapter 6 Ecology of E-PUB: DGGE
88
CHAPTER 6
Ecological Study of Environmental Pentose-Utilizing
Bacteria by Denaturing Gradient Gel Electrophoresis
(DGGE)
6.0 Summary
Previously in this thesis (chapter 3), the process of isolating pentose metabolizing bacteria from
a pentose rich environment was described, in order to assess their industrial potential. While
this autecological approach has many advantages, it must be recognized that the natural eco-
niche of these organisms is not a simple one, and that there is a vast amount of knowledge to be
gained from using a synecological approach, where the bacterial community of a site is
examined as a whole. This approach is best applied using metagenomic methods, such as the
DGGE technique employed here. This research analysed microbial communities in sugar mill
samples such as water, soil, and bagasse leachate collected from five cane-growing areas in
Queensland, Australia, using Denaturing Gradient Gel Electrophoresis (DGGE) and subsequent
DNA sequencing.
It was found that a variety of organisms were present in the bagasse leachate that was not
present in other samples. The most common environmental bacteria Brevibacillus brevis,
Rhodospirillaceae bacterium, Bacillus sp, Vibrio sp and Pseudomonas sp were present in all
samples. Corynebacterium was found in the soil of Proserpine, Mackay and Maryborough sugar
mills.
6.1 Introduction
The dynamics of bacterial communities within a hemicellulose-enriched environment represents
an opportunity for commercial enterprise. There is a wide variety of valuable end-products from
the natural microorganisms after the catabolism of lignocellulosic materials.
Applied molecular microbiology is a rapidly growing research area. One of the branches of this
discipline is involved in the development of molecular methods for the identification and
monitoring of microorganisms in natural ecosystems. The principal reason for the use of culture-
independent techniques is the lack of knowledge of the real conditions under which most
bacteria are growing in their natural habitat and the difficulty in developing growth media that
accurately re-create these conditions (Li et al., 2006). Molecular methods are also characterised
by rapidity and reliability. Genetic fingerprinting techniques are able to provide a profile
representing the genetic diversity of a microbial community from a specific environment

Chapter 6 Ecology of E-PUB: DGGE
89
(Ercolini, 2004). Denaturing gradient gel electrophoresis (DGGE) is now a commonly used
technique in molecular biology and has become a fundamental method of environmental
microbiology for the characterization of population structure and dynamics (Sanchez et al.,
2007; Green et al., 2007) . The method is powerful, and can rapidly provide a tangible
characterization of community diversity and composition, and shifts in population can be readily
demonstrated.The only major drawback is the inability to load all samples onto the same gel and
the potential for gel-to-gel variation, which influences DGGE analysis (Nunan et al., 2005). In
addition to its use in environmental analyses, DGGE is also used in the medical field for the
detection of mutations, including single nucleotide polymorphisms (SNPs) (Green et al., 2007).
16S rDNA gene sequencing is required to identify the individual microbial species from DGGE
analysis. 16S rDNA gene sequencing is a powerful tool that has been used to trace phylogenetic
relationships between bacteria, and to identify bacteria from various sources, such as
environmental sources (Neufeld et al., 2006) or clinical specimens (Clarridge, 2004 ). This
technology is used today in clinical laboratories for routine identification, especially for slow-
growing, unusual or fastidious bacteria, but also for bacteria that are poorly differentiated by
conventional methods (Clarridge, 2004; Vandamme et al., 1996).
Phenotypic methods present some inherent problems. There can be a substantial amount of
variability among strains belonging to the same species; the corresponding database may not yet
include newly described species; and the test may rely on an individual and subjective
interpretation (Raoult et al., 2004). Identification based on the 16S rDNA sequence is of interest
because the ribosomal small subunit (SSU) exists universally among bacteria and includes
regions with species-specific variability, which makes it possible to identify bacteria to the genus
or species level by comparison with databases in the public domain (Clarridge, 2004 ). The
molecular approach is important for species definition and identification (Clarridge, 2004 ;
Rosselló-Mora and Amann, 2001; Raoult et al., 2004). Other authors (Bosshard et al., 2003; Tang
et al., 2000) have also reported 16S rDNA sequencing use as a tool for bacterial identification.
They have usually compared this molecular identification tool favourably to classical phenotypic
methods. The great potential of the method has been reported for Gram-positive rods and
coryneform bacterial identification (Bosshard et al., 2004; Tang et al., 2000). This is of particular
interest in the present study, as these categories are the focus of this work.

Chapter 6 Ecology of E-PUB: DGGE
90
6.2 Materials and methods
6.2.1 Extraction of bacterial DNA from samples
Extraction of the DNA from soil and bagasse stockpiled samples was performed using the MOBIO
power soil DNA isolation kit (MOBIO Laboratories inc. USA). The MOBIO ultra clean water DNA
isolation kit was used for DNA extraction from liquid waste and bagasse leachate samples. All
sample details are provided in Table 6.1. Extracted DNA was quantified spectrophotometrically
(Beckman Coulter, Australia) by measuring the absorbance ratio A260:A280, which determines
the purity of the DNA.
Table 6.1: List of the samples collected from different sugar mills
Sample no. Sample nature Sample name Location Sample code
1 Liquid Floor dump sump Proserpine FDS-P
2 Liquid Cooling tower water Proserpine CTW-P
3 Liquid Floor dump sump Mackay FDS-MK
4 Liquid Cooling tower water Mackay CTW-MK
5 Liquid Floor dump sump Maryborough FDS-M
6 Liquid Cooling tower water Maryborough CTW-M
7 Liquid Bagasse leachate Mackay BGL-MK
8 Solid Mud sample Proserpine MS-P
9 Solid Decaying trash from cane farm Proserpine DT-P
10 Solid Bagasse from stockpile Proserpine BG-P
11 Solid Mud sample Maryborough MS-M
12 Solid Decaying trash from cane farm Maryborough DT-M
13 Solid Bagasse from stockpile Maryborough BG-M
14 Solid Mud sample Mackay MS-MK
15 Solid 46 Caswells soils Mackay CS-MK
16 Solid 42 Blair soil Mackay BS-MK
17 Solid 58 Effluent pond soil Mackay EPS-MK
18 Solid 55 Tropic Isle soil Mackay TIS-MK

Chapter 6 Ecology of E-PUB: DGGE
91
6.2.2 Sample description
The Floor dump sump (FDS) holds the liquid waste from washing the raw materials, it is filtered,
passes through the pipe network, and finally is stored in the sump; cooling tower water (CTW).
Figure 6.1: Typical sugar processing mill. Sampling sites marked with arrows. (modified from:
http://www.enterprisewhitsundays.com.au/index.php/download_file/view/109/.)

Chapter 6 Ecology of E-PUB: DGGE
92
Bagasse is sugar cane waste after pulping and extraction; bagasse leachate is liquid in nature,
created by rainwater passing through the bagasse heaps from the bagasse stockpile. All bagasse
piles are covered with tarps and secured with strapping/tyres. All pads are constructed with
road base, self draining to leachate catchment system. A storm water leachate collection system
was collection from each of the storage sites or gravity run-off to mill effluent systems. Water
collected in leachate collection dams/ponds pumped to mill effluent systems.
The sampling site and a diagram of the sugar processing are shown in Figure 6.1. The strategy for
selecting these sites was to ensure samples that might be expected to have hemicellulose-
enrichment. However, cooling tower water (CTW) has no physical connection to the sugar
waste. For this reason, CTW was selected as a control, non-enriched sample. A major objective
of this study was to compare the microbial diversity within hemicellulose-enriched and non-
hemicellulose-enriched habitats. This was because work continues on the bagasse stockpiled
area with respect to mitigating environmental and health issues, associated with large scale
bagasse storage (Womersley, 2006). Leachate and contaminated stormwater from bagasse
stockpiled are generally confined to drains and ponds. Odour from the bagasse stockpile is an
issue, and may include the formation of acetic acid in older piles. Most odour problems are
found at the pile breakdown point (site, and also time). Odour issues persist mainly on site
where bagasse may be confined to conveying systems for extended periods of time.
6.2.3 16S rDNA PCR amplification
PCR amplification was performed in a 25-μL (total volume) reaction mixture. The final
concentration of different components in the mixture was: 50 ng of purified DNA, 0.4 μM of
each primer (Sigma-Aldrich), 200 μM of each deoxynucleoside triphosphate (dNTP, Roche), 1.5
mM MgCl2 (Invitrogen), 1× thermophilic DNA polymerase, PCR 10×reaction buffer (Roche), 1.25
U per 50 μL of Taq DNA polymerase (Roche), and DNAse and RNAse free filter sterilized water
(GIBCO ultrapure water-Invitrogen). PCR amplification of 16S rDNA bacterial genes was carried
out using the universal primers (forward primer 5′ GAGTTTGATCCTGGCTCAG 3′ and reverse
primer 5′ ACGGCTACCTTGTTACGACTT 3′). PCR was performed using the following cycle
conditions: 95°C for 5 min followed by 25 cycles of 94°C for 1 min per cycle, 55°C for 1 min, 72°C
for 1 min and a final extension step of 72°C for 10 min. The PCR samples were amplified using a
PCR thermal cycler (Bio-Rad, DNA engine, Peltier, Thermal cycler, USA) and PCR products were
visualized on a 1% agarose gel stained with ethidium bromide.

Chapter 6 Ecology of E-PUB: DGGE
93
6.2.4 Denaturing gradient gel electrophoresis (DGGE)
Duplicate DGGE was performed with the Bio-Rad DCode System (Bio-Rad, Australia). Equal
volumes of each PCR product were loaded onto polyacrylamide gels with a denaturing gradient
of 30% (7% (w/v) acrylamide–bisacrylamide (37.5:1), 2.55 M urea, 14.68% formamide) to 70%
(7% (w/v) acrylamide–bisacrylamide (37.5:1), 3.57 M urea, 20.56% formamide) with Tris-acetate
EDTA buffer (pH 7.4), at 65° and 60 V, for 16 h. After electrophoresis, the gels were soaked for
30 min in gel-red nucleic acid gel stain (1:10000 dilution) (BIio-Rad, Australia). The stained gels
were immediately photographed on a UV transilluminator (Bio-Rad, Australia).
6.2.5. Analysis of DGGE patterns
The DGGE patterns were analysed using the Bio-Rad software to create a phylogenetic tree
based on the molecular weights of each visible band on the DGGE gel. The gel image was
imported into Total Lab 120 software (Nonlinear Dynamics Ltd, United Kingdom) to determine
the band patterns for each sample. Band location and intensity were imported into PrimerE
software (Plymouth Marine Labs, United Kingdom) and a similarity matrix was created using the
Bray-Curtis method, to observe the similarity of community fingerprints. The Bray-Curtis
similarity coefficient was used to determine site similarities based on organism abundances. It
reflects differences between two samples due both to a differing community composition
and/or a differing total abundance.
6.2.6 Re-extraction of DNA from DGGE gels followed by DNA sequencing
Bands selected for sequence analysis were carefully excised from the DGGE gel with a sterile
scalpel. The middle portion (avoiding edges) of each band was selected to minimize the DNA
contamination and to obtain high concentrations of DNA. The slices were placed in 2-mL
sterilized microfuge tubes. DNA was purified using the MOBIO (MOBIO laboratories, USA) gel
extraction kit. The DNA was quantified by the spectrophotometric method at 260/280 nm. A 5
µL sample of the eluate was used as template DNA for a PCR performed with the universal 16S
rDNA primers and the conditions described above for environmental samples (section 6.2.3).
Five µL of each PCR product was subjected to agarose gel electrophoresis to check product
recovery and to estimate product concentration.
6.2.7 16S rDNA gene sequencing methodology
16S rDNA sequencing was performed using a Dye Terminator Cycle Sequencing reaction method
that included 200 ng PCR product, 3.2 pmol primer, 1 μL big dye, 3.5 μL sequence buffer and
water in a total volume of 20 μL. PCR was performed using a DNA engine, (Peltier, Thermal

Chapter 6 Ecology of E-PUB: DGGE
94
cycler) programmed for using forward and reverse sequencing primers 16S F 5′
GAGTTTGATCCTGGCTCAG 3′ and R 5′ ACGGCTACCTTGTTACGACTT 3′ respectively. The
sequencing reaction was thermally cycled as follows on the DNA Engine (Peltier, Thermal cycler):
95 °C for 1 min (1 cycle); 98 °C for 45 s, 50 °C for 10 s, 60 °C for 4 min (1 cycle); 98 °C for 15 s, 50
°C for 10 s, 60 °C for 4 min (29 cycles); 4 °C for 5 min (1 cycle); 10 °C hold.
Sequencing products, unincorporating terminators and primers were purified with a 70%
ethanol wash, using an ethanol/EDTA method and big dye x terminator purification kit according
to the Griffith University, Brisbane, Australia DNA sequencing protocol
(http://www.griffith.edu.au/science-aviation/dna-sequencing-facility/services/dna-
sequencing/sequencing-protocolm ). Sequencing products were analysed using an ABI 3500
Sequencer (Applied Biosystems). Consensus sequences were assembled from the forward and
reverse sequences and edited with Sequencer software (Bio- Edit). Consensus results were
compared with sequences in GenBank, using the BLAST sequence similarity search.
6.2.8 Statistical analysis
Statistical analysis of band patterns was performed using the relative band intensity within a
lane as well as the presence vs. absence species. Cluster diagrams were created using the
unweighted pair group method with arithmetic mean (UPGMA). The Bray-Curtis similarity
coefficient was used whereby the relative intensity of each band was also taken into account. In
addition, non-metric multidimensional scaling (MDS) analysis was done in parallel to further
elucidate the differences in DGGE profiles.
Analysis of the bacterial community DGGE profiles was performed with the Bio-Rad software
package according to the provider's instructions. The DGGE gel photographs were screened for
the presence (1) or absence (0) of bands on the DGGE gel. The unweighted pair-group method
with arithmetic averages (UPGMA) algorithm was used (as part of the analysis software), for the
construction of phylogenetic dendrograms.
6.3 Results
6.3.1 PCR–DGGE analysis
The DGGE profiles of bacterial communities in seven liquid and 11 solid samples from
Queensland sugar mills were observed. Lane numbers 1 to 6 contained the extracts from liquid
samples; lane seven contained the bagasse leachate (BGL-MK) and lanes eight to 18 contained
the solid samples.

Chapter 6 Ecology of E-PUB: DGGE
95
6.3.1.1 Analysis of the microbial community structure using the Bray-Curtis method
All microbial communities of solid and liquid samples from the Proserpine, Mackay and
Maryborough sites remained clustered within their respective treatments (Figure 6.2). The Bray-
Curtis dendrogram (Figure 6.2) demonstrated that the association was strongest among
communities in the solid samples in which eight replicate plots shared a 65% similarity.
Additionally, four groups of liquid samples also shared a 65% similarity among plots, except the
cooling tower water from Maryborough and Proserpine. These two samples shared only a 50%
similarity. The bagasse leachate (Mackay only) samples also showed a 65% similarity. However
BG-P shared about 80% similarity with the remainder of the samples (Figure 6.2). The liquid
samples (floor dump sump, cooling tower water and bagasse leachate as available from the
various mills) are also analysed on the MDS plot in Figure 6.5. This further examination was
performed in order to better understand the microbial communities of the liquid samples from
Figure 6.2. Although bagasse form Proserpine and Maryborough were not liquid samples, they
were included in Figure 6.5 to demonstrate the difference between bagasse, and bagasse
leachate, which is considerable.

Chapter 6 Ecology of E-PUB: DGGE
96
Figure 6.2: Community fingerprint analysis using the Bray-Curtis method.
The sample codes are represented in detail in Table 6.1 (Sample type – Mill location).
6.3.1.2 Analysis of microbial community structure in liquid samples
In this part of the study, liquid samples were analysed separately and Figure 6.3 shows the DGGE
profiles of the microbial communities of seven liquid samples including BGL-MK.
FDS-P (1)
CTW-P (2)
FDS-MK (3)
BGL-MK (7)
MS-P (8)
DT-P (9)
CTW-M (6)
CTW-MK (4)
FDS-M (5)
BG-M (13)
DT-M (12)
MS-M (11)
BG-P (10)
MS-MK (14)
CS-MK (15)
BS-MK (16)
EPS-MK (17)
TIS-MK (18)

Chapter 6 Ecology of E-PUB: DGGE
97
Figure 6.3: DGGE profiles of bacterial community structures in liquid samples. Lanes one to seven represent the sample codes FDS-P, CTW-P, FDS-MK, CTW-MK, FDS-M, CTW-M and BGL-MK respectively.
Figure 6.4: Phylogenetic tree analysis of liquid samples by Unweighted Pair Group Method with
Arithmetic Mean (UPGMA)
FDS-P
CTW-P
CTW-MK
CTW-M
FDS-MK
FDS-M
BGL-MK

Chapter 6 Ecology of E-PUB: DGGE
98
Figure 6.4 represents the DGGE gel profiles that were incorporated into a phylotree by using an
unweighted pair-group average (UPGMA) analysis. UPGMA analysis joins clusters based on the
average distance between all members in the two groups. This analysis revealed that the DGGE
band patterns for each site clustered separately. The greatest difference was found between the
profiles of sample #7 (bagasse leachate from Mackay) and all other liquid samples.
The bagasse leachate sample population (BGL-MK - band 7) shared only 61% similarity with that
of all other liquid samples. On the other hand, a 90 % similarity was noted between floor dump
sump sample populations at Proserpine and Mackay (FDS-P – lane 1 and FDS-MK - lane3).
Interestingly, populations from the cooling tower water at Mackay (CTW-MK – lane 5) and the
floor dump sump from Maryborough (FDS-M - lane 4) have more than an 80% similarity;
whereas the populations of the same category of samples from different locations (CTW-P – lane
2 and CTW-M – lane 6) show only 73% similarity with most liquid samples, with the exception of
bagasse leachate.
Whilst the multidimensional scaling plots of DGGE patterns revealed different patterns of
bacterial communities compared to the source of the sampling (Figure 6.2), in most cases no
clear trend could be observed. This method demonstrated the separation of samples by sample
type but showed minimal effect of location. A variety of types of liquid samples from different
sources were further analysed and the results are displayed in Figure 6.5. From this analysis,
bagasse leachate and bagasse from stockpiled could only be compared with similar samples.
Figure 6.5 is a MDS analysis of phylogenetic relationships (from Figures 6.3 and 6.4) of liquid
samples (FDS-P, CTW-P, FDS-MK, CTW-MK, FDS-M, CTW-M and BGL-MK) including bagasse
samples BG-P and BG-M.
The MDS analysis of these patterns revealed that the structure of the microbial community from
liquid samples; FDS-M, CTW-M, FDS-P, FDS-MK and CTW-MK; were not significantly different,
although CTW from Proserpine mill was slightly different from the other sites (Figure 6.5). In
contrast, bagasse leachate from Mackay (BGL-MK) was significantly different to all other
samples. Also as seen in Figure 6.5, no significant differences were observed between the two
bagasse samples (BG-P, BG-M) from the Proserpine and Maryborough sugar mills.

Chapter 6 Ecology of E-PUB: DGGE
99
Figure 6.5: Non-metric multi-dimensional scaling (MDS) plot analysis of DGGE bands for liquid
samples plus two bagasse samples
6.3.1.3 Analysis of microbial community structure in solid samples
Each of the distinguishable bands in the separation pattern represents an individual bacterial
species. Clustering of the profiles revealed that there were very large differences between the
profiles of the solid samples. The profiles of solid samples MS-P, DT-PMS-M, DT-M, MS-MK, CS-
MK, EPS-MK, and TIS-MK had showed (Figure 6.7) 83% - 88% similarity with respect to their
clustering. The profile of bagasse from Proserpine (BG-P) was quite different and separate from
other profiles and belonged to a single group, with a similarity of 54% to the profile of sample
BS-MK and others.

Chapter 6 Ecology of E-PUB: DGGE
100
Figure 6.6: DGGE profiles of bacterial community structures in soil samples
Figure 6.7: Phylogenetic tree analysis of solid samples. All the abbreviations of the sample codes
are listed in Table 6.1.
MS-P
CS-MK
MS-MK
BG-M
DT-M
MS-M
BG-P
DT-P
BS-MK
EPS-MK
TIS-MK

Chapter 6 Ecology of E-PUB: DGGE
101
There was about 71% similarity between the two groups. Group (a) consisted of samples MS-
MK, CS-MK, EPS-MK, BS-MK, and group (b) of samples MS-M, DT-M, BG-M). This analysis
indicated the bacterial diversity of the hemicellulose-enriched soil waste of Queensland sugar
mills.
6.3.2 16S rDNA sequencing analysis of selected DGGE bands
The 17 sequences were compared with by BLAST software in the NCBI database (Figure 6.8).
Only partial sequences were aligned and the similarity between these sequences of
microorganisms in Genbank is shown in Table 6.2.
Figure 6.8: DGGE gel image indicating the lane numbers
Lanes 1 to 18 represent the sample codes FDS-P, CTW-P,FDS-MK,CTW-MK,FDS-M,CTW-M, BGL-
MK, MS-P, DT-P, BG-P, MS-M, DT-M, BG-M, MS-MK, CS-MK, BS-MK, EPS-MK, TIS-MK
respectively.
Using a threshold of greater than 65%-99% similarity for positive identification, evidence was
found for the presence of Brevibacillus brevis, Rhodospirillaceae bacterium, Rhodospirillaceae
bacterium, Corynebacterium sp., Pseudomonus fluorescens, Bacillus subtilits, endophytic
bacteria, Pseudomonas aeruginosa, Vibrio vulnificus, Azospirillum brasilense, Roseomonas
fauriae, Bulkholderia cepacia, Bacillus cereus, Bacillus thuringiensis, Alcaligenes faecalis, Vibrio
ichtyoenter and Vibrio aestuarianus in all samples tested (Table 6.2 and 6.3).

Chapter 6 Ecology of E-PUB: DGGE
102
Table 6.2: 16S rDNA sequencing results including samples and DGGE band details
Lane no
Sample nature
Sample name Band No
Species
1 Liquid Floor dump sump from Proserpine (FDS-P)
1,2,5, 10,17
Brevibacillus brevis, Rhodospirillaceae sp., Bacillus subtilits, Roseomonas fauriae, Vibrio aestuarianus
2 Cooling Tower water from Proserpine(CTW-P)
1,2,5, 15,17
Brevibacillus brevis,Rhodospirillaceae sp., Bacillus subtilits, Vibrio ichtyoenter, Vibrio aestuarianus,
3 Floor dump sump from Mackay (FDS-MK)
1,2,5, 9,11,17
Brevibacillus brevis, Rhodospirillaceae sp., Bacillus subtilits, Azospirillum brasilense, Burkholderia cepacia, Vibrio aestuarianus
4 Cooling Tower water from Mackay (CTW-MK)
1,2,6, 17
Brevibacillus brevis, Rhodospirillaceae sp., Endophytic bacterium, Vibrio aestuarianus
5 Floor dump sump from Maryborough(FDS-M)
1,2,4,5, 17
Brevibacillus brevis, Rhodospirillaceae sp., Pseudomonas fluorescens, Bacillus subtilits, Vibrio aestuarianus
6 Cooling Tower water from Maryborough (CTW-M)
1,2,7, 8,12,17
Brevibacillus brevis, Rhodospirillaceae sp., Pseudomonas aeruginosa, Vibrio vulnificus, Bacillus cereus, Vibrio aestuarianus
7 Bagasse leachate from Mackay (BGL-MK)
1,5,8, 13,14,17
Brevibacillus brevis, Bacillus subtilits, Vibrio vulnificus, Bacillus thuringiensis, Alcaligenes faecalis, Vibrio aestuarianus
8 Solid Mud sample from Proserpine (MS-P)
1,4,5,6,8
Brevibacillus brevis, Pseudomonas fluorescens, Bacillus subtilits, Endophytic bacterium, Vibrio vulnificus
9 Decaying trash from cane farm Proserpine( DT-P)
1,2,4,5, 17
Brevibacillus brevis, Rhodospirillaceae sp., Pseudomonas fluorescens, Bacillus subtilits, Vibrio aestuarianus
10 Bagasse from Proserpine(BG-P)
1,3,4,5,8, 17
Brevibacillus brevis, Corynebacterium sp., Pseudomonas fluorescens, Bacillus subtilits, Vibrio vulnificus, Vibrio aestuarianus
11 Mud sample from Maryborough (MS-M)
1,3,4,5, 11,17
Brevibacillus brevis, Corynebacterium sp., Pseudomonas fluorescens, Bacillus subtilits, Burkholderia cepacia, Vibrio aestuarianus
12 Decaying trash from cane farm Maryborough(DT-M)
1,3,4,5, 12,14,15,17
Brevibacillus brevis, Corynebacterium sp., Pseudomonas fluorescens, Bacillus subtilits, Bacillus cereus, Alcaligenes faecalis, Vibrio ichtyoenter, Vibrio aestuarianus
13 Bagasse from Maryborough (BG-M)
1,3,4,5, 13,17
Brevibacillus brevis, Corynebacterium sp., Pseudomonas fluorescens, Bacillus subtilits, Bacillus thuringiensis, Vibrio aestuarianus
14 Mud sample from Mackay (MS-MK)
1,3,5,7, 8,10,17
Brevibacillus brevis, Corynebacterium sp. Bacillus subtilits, Pseudomonas aeruginosa, Vibrio vulnificus, Roseomonas fauriae, Vibrio aestuarianus
15 46 caswells soils from Mackay (CS-MK)
1,3,5,8, 14,15,17
Brevibacillus brevis, Corynebacterium sp., Bacillus subtilits, Vibrio vulnificus, Alcaligenes faecalis, Vibrio ichtyoenter, Vibrio aestuarianus
16 46 Blair soil from Mackay (BS-MK)
1,4,5,12, 13,16
Brevibacillus brevis, Pseudomonas fluorescens, Bacillus subtilits, Bacillus cereus, Bacillus thuringiensis, β- proteobacterium,
17 58 Effluent pond from Mackay (EPS-MK)
1,4,5,8, 16,17
Brevibacillus brevis, Pseudomonas fluorescens, Bacillus subtilits, Vibrio vulnificus, β-proteobacterium sp., Vibrio aestuarianus
18 55 Tropic Isle Mackay (TIS-MK)
1,2,3 Brevibacillus brevis, Rhodospirillaceae sp., Corynebacterium sp.,

Chapter 6 Ecology of E-PUB: DGGE
103
Table 6.3: 16S rDNA sequencing result with closest matched Genus/species based on the
percentage (%) of similarity
Closest matched genus/species using
the Universal Primer 16S rDNA
Sequence similarity in
% (no. of bases)
Taxonomic group
Brevibacillus brevis 93% Proteobacteria
Uncultured Rhodospirillaceae sp., 95% Proteobacteria
Uncultured Corynebacterium sp 96% Actinobacteria
Pseudomonas fluorescens, 97% Proteobacteria
Bacillus subtilits 95% Firmicutes
Endophytic bacterium 70%
Pseudomonas aeruginosa 98% Proteobacteria
Vibrio vulnificus 93% Proteobacteria
Azospirillum brasilense, 95% Proteobacteria
Roseomonas fauriae 95% Proteobacteria
Burkholderia cepacia 97% Proteobacteria
Bacillus cereus 99% Firmicutes
Bacillus thuringiensis 99% Firmicutes
Alcaligenes faecalis 93% Proteobacteria
Vibrio ichtyoenter 92% Proteobacteria
β- proteobacterium 66% Proteobacteria
Vibrio aestuarianus 94% Proteobacteria
6.4 Discussion
Previous research has shown that PCR-DGGE provides detailed information about changes in and
diversity of microbial community structures in the environment, compared to plate count
methods (Li et al., 2006). The results of this study demonstrate that DGGE and 16S rDNA
sequencing used together provide insight into the bacterial community composition of solid and
liquid samples and the different genotypic patterns for each of the bacterial communities that
were observed by these means. To the best of our knowledge, very limited research on the
identification of the ecological diversity in hemicellulose-enriched sites has been conducted.

Chapter 6 Ecology of E-PUB: DGGE
104
In this study, microbial DNA was analysed from solid (soil, bagasse, decaying trash) and liquid
(bagasse leachate, floor dump sump) sources by the DGGE method. The DGGE of PCR-amplified
16S rRNA gene fragments clearly demonstrated the microbial characteristics of environmental
(solid and liquid) samples from sugar mills in Queensland.
There was a couple of difficulties encountered in this phase of the research. Unfortunately only
one bagasse leachate sample was available, due to a drought at the time of the sampling. The
Mackay sample was sent later in the season, after rainfall events. Another drawback was the
difficulty in loading all samples onto the same gel and the resulting potential for gel-to-gel
variation, which influences DGGE analysis (Nunan et al., 2005). The results were also limited by
the use of Universal primers in the PCR.
Our main aim was to analyze microbial diversity in hemicellulose-enriched habitats such as
bagasse leachate. DGGE profiles of bacterial communities in six liquid waste samples, one
bagasse leachate, two bagasse stockpile samples and nine soil samples from Queensland sugar
mills were observed. Each sample was analyzed in duplicate with the same result. Many equally
intense bands, indicating the presence of a large number of equally abundant ribotypes, were
observed for all sample types.
The DGGE profiles of liquid samples revealed that the band patterns for cooling tower water
from Mackay (CTW-MK) and Proserpine mill (CTW-P) were found to be most similar to each
other, while bagasse stockpiled from the Proserpine mill (BG-P) had the least similarity with the
patterns of the other solid samples. From the MDS analysis, it could be demonstrated that the
sugar cane stored in the floor before crushing, does not contain much freely available
hemicellulose, which may be the reason why there were no significant differences between the
microbial populations found in the cooling tower water (CTW) and floor dump sump (FDS).
Alternatively, the cooing tower water is more ubiquitous than anticipated, and other sampling
sites may be been contaminated by the populations from the cooling tower. This is feasible, as
cooling towers generally produce a fine, but widespread spray of water for a considerable
distance across an industrial site. Thus, the microbial population of the cooling tower may have
been transported to other parts of the site.
Bagasse leachate is expected to have a high hemicellulosic composition compared to that of
stockpiled bagasse, due to the way that it is produced, by slow penetration of rainwater through
a bagasse pile, gradually accumulating soluble products, such that the end result is quite
concentrated. This may be the reason that the DGGE pattern for bagasse leachate (BGL) was
significantly different to those of the stockpiled bagasse, which was also significantly different to

Chapter 6 Ecology of E-PUB: DGGE
105
cooling tower and floor dump samples. Bagasse leachate from Mackay (BGL-MK) and bagasse
stockpiled from Maryborough mill (BG-M) had a 76% similarity in population, probably because
of the similar nutritional composition.
Factors other than hemi-cellulose concentration in the samples, that may potentially contribute
to the variation in microbial communities between samples, include: temperature (ambient cf
factory floor); moisture levels (ranging from dry to very moist) and the related factor of relative
humidity; oxygen availability (either natural or resulting from microbial activity); soil type (clay,
sandy, humous) of the region; and even, possibly, the time of day that sampling occurred (which
may have influenced several of the mentioned factors). An example of the type of habitat likely
to be encountered is freshly baled bagasse, which contains 50 % water and about 3 % sugar, and
heats rapidly after stacking (Lacey, I97I). The temperature remains above 40°C for few weeks
with a maximum of 50°C to 60°C, providing good growing conditions for thermophilic
actinomycetes. Unfortunately, none of these factors were recorded for this project, so it is not
possible to calculate their contribution to the variation between populations in various samples.
According to the phylogenetic tree analysis, the microbial population in bagasse leachate from
Mackay was the most divergent among all of the liquid samples. The floor dump sump from
Proserpine and Mackay sugar mills had the highest similarity (90%) between their microbial
populations probably due to the similarity of the nature of the samples. The floor dump sump
and cooling tower water from Maryborough had more than 80% similarity possibly because both
samples were collected from the same location (Maryborough sugar mill). The remainder of the
samples were highly diverse.
Bagasse leachate, only available from the Mackay mill, was quite different to other liquid
samples, and in fact showed more similarity with the soil samples from the Mackay region. This
indicated that the bacterial communities were similar on the basis of geographical location.
The alkaline-tolerant bacteria Alcaligenes faecalis was found in bagasse leachate and caswells
soils from Mackay mill (CS-MK). The reason may be peculiar to the area (Mackay), such as an
effect of soil chemistry or climate near that sugar mill. Also nitrogen fixing bacteria Azospirillum
brasilense was found in the FDS-Mackay sample. Nitrogen-fixing β-proteobacteria was also
found in Mackay sugar mill. The β-proteobacteria consist of several groups of aerobic or
facultative bacteria, which are often highly versatile in their degradation capacities and play a
role in nitrogen fixation in various types of plants, oxidizing ammonium to produce nitrite, an
important chemical for plant function.

Chapter 6 Ecology of E-PUB: DGGE
106
Since many endospore-forming species can effectively degrade a series of biopolymers such as
proteins, starch, pectin, cellulose, they are assumed to play a significant role in the biological
cycles of carbon and nitrogen. From soil, by direct contact or air-borne dust, endospores can
contaminate almost anything that is not maintained in a sterile environment. They may play a
biodegradative role in whatever they contaminate, and thereby they may be agents of
decomposition and decay. Several Bacillus species are especially important as food spoilage
organisms (Zavarzin, 2006). These abilities and roles are very relevant to the industrial
application that the current research was intended to explore.
All soil samples (MS-M, DT-P; DT-P, TIS-MK; MS-MK, CS-MK; MS-P, EPS-MK) among the solid
samples had more than 80% similarity, but this did not apply to the two bagasse stockpiled
samples. Bagasse stockpiled from Proserpine mill (BG-P) and Maryborough sugar mill (BG-M)
were highly divergent populations compared to the soil samples. The possible reason of this
finding may be the similar composition of nutritional elements present in microbial habitat. Also
microbial growth factors (aeration, pH, RH, temperature, carbon sources) may influence the
source, producing a divergent microbial population.
Corynebacterium spp. were isolated (chapter 3) previously from the soil samples of sugar mills.
Similarly, Corynebacterium spp. were found in this study in solid samples (mud, decaying trash,
and bagasse stockpiled) from Proserpine, Mackay and Maryborough regions, but not from any of
the liquid samples. Corynebacterium spp. that are capable of producing amino acids such as
glutamic acid, valine, isoleucine, threonine, aspartic acid and alanine (Hermann, 2003) were
found in most soil samples. This finding revealed that the presence of commercially valuable
Corynebacterium spp. in sugar industry-related solid samples is widespread.
Most of the microorganisms or groups of microorganisms found in the samples were those
commonly expected in soil and water samples. However, only the soil samples contained the
coryneform group that includes the isolates reported in Chapter 3, and the subject of the
research conducted. It is conjectured that the water samples that were subjected to DGGE
analysis had not undergone any enrichment process, and so would have been colonised mainly
by sucrose-utilizing organisms, that being the carbon source most commonly available in the
selected sites and samples.
The most pentose-rich samples were expected to be the bagasse and bagasse leachate, because
of the close association with sugar cane refuse, and these certainly had a different community
structure to the other samples. However, the bagasse leachate sample also did not contain
detectable levels of Corynebacterium spp. in the DGGE analysis. The samples used in the original,

Chapter 6 Ecology of E-PUB: DGGE
107
culture-based isolation procedure (Chapter 3) were also soil samples. So it must be concluded
that the target coryneform micro-organisms were simply not able to grow in water, at least in
the environments that were under investigation. It is known that Corynebacterium spp. may be
found in both water and soil (Mallory et al., 1977), just as it is also known that members of this
genus, some of which have now been reassigned to other genera (eg. Rhodococcus equii), are
able to cause plant and animal diseases as a result of contact with contaminated soil and water
(Takai et al., 1986).
It is not clear why the Corynebacterium genus was not present in the sampled waters, but clearly
present in the soil. One explanation may be that the drought that was affecting the area when
the samples were first taken, may have selected for soil coryneforms by virtue of the dryness of
the environment. The liquid samples, with the exception of bagasse leachate, would probably
have been originally source from treated water, before being pumped around the site, where
they became contaminated by indigenous microflora. The treatment of the water would have
negatively influenced the ability of the Corynebacterium spp. to grow.
There are some bacteria found in common to both solid and liquid samples. Phylogenetically,
the sequences were distributed into five groups: Bacillus spp, nitrogen fixing Proteobacterium,
Vibrio spp, Pseudomonas sp and Corynebacterium spp. The dominant bands of all of the samples
corresponded to the β-Proteobacterium group, while the sequences recovered from bagasse
leachate samples belonged to Pseudomonas spp. and coryneform groups. Based on the
sequences of the bands excised from each of the samples, there were approximately five to six
different characteristic bands present in each of the liquid and solid samples, the dominant
groups being Proteobacteria and nitrogen-fixing bacteria. It was also found that the presence of
Gram-negative bacteria and Gram-positive bacteria was equally distributed among samples.
The presence of Vibrio spp in these samples would once have been thought to be quite unusual
(Colwell et al., 1992) however current knowledge is that this genus may be found in soil and dust
and is often able to degrade chitin. In addition, this bacterium is known to have the ability to
degrade hemicellulose from the soil along with other genera found in the samples, such as
Bacillus spp., Pseudomonas spp., and Erwinia (My Agriculture Information Bank/Soil
Microbiology). It would be expected that samples from environments surrounding and within
sugar cane mills would include those genera capable of degrading such materials as: cellulose
(Pseudomonas, Cytophaya, Spirillum, Actinomycetes and Cellulomonas); Lignin (Pseudomonas,
Micrococcus, Flavobacterium, Zanthomonas and Streptomyces); and Pectin (Erwinia) (My
Agriculture Information Bank/Soil Microbiology).

Chapter 6 Ecology of E-PUB: DGGE
108
Brevibacillus brevis, Rhodospirillaceae bacterium, Bacillus sp, Vibrio sp and Pseudomonas were
the most wide-spread microorganisms, as they were found in all samples. These microbes are
decomposers, which have the ability to recycle nutrients. Microorganisms have a special role as
degraders in the biogeochemical cycles.
Such microorganisms, because of their ability to degrade a wide range of substrates, are often
used for in-situ microbial biodegradation or bioremediation of domestic, agricultural and
industrial wastes and subsurface pollution in soils, sediments and marine environments. The
ability of each microorganism to degrade toxic waste depends on the nature of the
contaminants. Since most sites typically have multiple pollutant types, the most effective
approach to microbial biodegradation is to use a mixture of bacterial species and strains, each
specific to the biodegradation of one or more types of contaminants. The composition of the
indigenous and added bacteria is carefully controlled, in order to evaluate the activity level and
to permit modifications of the nutrients and other conditions for optimizing the bioremediation
process (Zavarzin, 2006; Koukkou, 2011; Watanabe and Kasai, 2008; Okabe and Kamagata,
2010).
Pseudomonas species, present in many of the samples, are amongst the toughest, most non-
fastidious and denitrifying of organisms. They are used for bioremediation purposes to clean up
oil spills; they grow on plastic surfaces, in disinfectants, and other difficult sites. They are
indigenous in water and can use almost any carbon source available (Madigan and Martinko,
2005). So the presence of Pseudomonas species is quite common in environmental water and
soil samples, and in this context they are known to break down a wide range of plant material,
including cellulose, hemicelluloses, lignin and proteins (My Agriculture Information Bank/Soil
Microbiology).
Brevibacillus brevis, Rhodospirillaceae bacterium, Bacillus sp, Vibrio sp and Pseudomonas play an
important role in the microbial ecosystem in the sugar industry. Large amounts of protein-
producing Brevibacillus brevis (Yamada et al., 1981) were present in all of the samples including
soil and liquid. The photosynthetic bacterium Rhodospirillaceae spp. was present in all liquid
samples except bagasse leachate.
According to Lacey (I97I), a new species Thermoactinomyces sacchari has been isolated from
sugar cane bagasse. Their heat-resistant spores contained dipicolinic acid which causes
bagassosis. Bagassosis is a respiratory disease caused by inhaling dust from mouldy, self-heated,
crushed sugar cane (bagasse). T. sacchari was isolated from the surface of freshly harvested

Chapter 6 Ecology of E-PUB: DGGE
109
sugar cane, and from muds from filter presses at the sugar mills, but was most abundant in
mouldy self-heated bagasse Other actinomycetes may also cause similar diseases. The
properties and structure of the spores of T. sacchari resemble spore-forming bacteria of the
genera Bacillus and Clostridium (Lacey, I97I). Regular microbial ecological analysis is needed to
monitor the risk of the health issues for people near sugar mill. It is possible that DGGE analysis
of the bagasse stockpiles would provide a rapid means of alerting the mill staff to dangerous
levels of such opportunistic pathogens.
It is apparent that the physical and nutritional characteristics of the microbial habitat have
impacted on the diversity of microbial communities supported. The identification of microbial
species capable of degrading hemicelluloses, has contributed to our research in support of the
aim of discovering species in this environment that have potential for industrial exploitation.
Although the DGGE analysis of the sugar mill samples did not clearly demonstrate the presence
of actinomycetes, DGGE analysis did reveal the presence of Corynebacterium spp. in particular.
Amongst the cultures isolated and characterized previously (Chapter 3), only some were found
to be present in sugar mill samples analysed by DGGE. Thus, some correlation between the
culture-based analysis of the sugar mill samples and DGGE profiles analysis was demonstrated in
this study. However, DGGE analysis provided a more detailed metagenomic picture than culture-
based methods had, and enabled the specific detection of potential pentose-utilizing microbes in
these samples.
6.5 Conclusions
Bacterial DGGE profiles generated using universal bacterial primers revealed the structural
composition of communities in liquid and solid samples. The most complex DGGE pattern
indicated the presence of a number of different bacterial taxa. DNA sequencing identification of
dominant members in DGGE profiles can aid in the selection of suitable isolation media and
conditions, since the phylogenetic positions of bacteria are often consistent with their
physiological properties and culture requirements (Throbäck et al., 2004).
Fingerprinting techniques such as DGGE provide separation of bacterial taxa based on 16S rRNA
banding patterns. In order to recognize the specific taxa that are shown as different bands, 16S
rDNA sequencing is also required. Bacterial 16S rDNA genes were readily detected and a variety
of microbial species from the sugar treatment waste were identified. While the results were
somewhat limited by the universality of the primers used in the PCR (Yu et al., 2008), the results
showed some interesting relationships. Brevibacillus brevis and Vibrio spp were found most
commonly in all of the samples. Rhodospirillaceae sp was found in all liquid samples except

Chapter 6 Ecology of E-PUB: DGGE
110
bagasse leachate. Rhodospirillaceae sp. was also found in DT-P and TIS-MK, different samples
from different locations. Alcaligenes faecalis was present in bagasse leachate and also in soils
from Mackay possible cause of the soil chemistry or climate of sugar mill. However, Burkholderia
cepacia was found as a minor microorganism in floor dump sump in Mackay and a mud sample
from Maryborough sugar mills, two quite different sites. It is important not to over-analyse
these minor taxonomic issues, as the population at this level of single genera may prove to be
ephemeral.
Most importantly, according to the DGGE profiles, Corynebacterium were found in the bagasse
stockpiled and soil samples of Proserpine, Mackay and Maryborough sugar mills. In Chapter 3,
Corynebacteria were reported as being isolated from Maryborough sugar mill’s soils. This
research discovered that Corynebacteria were found as a common bacterium in DGGE
fingerprint but only in soils, and were also found by traditional cultured methods. In addition,
bagasse leachate and bagasse stockpile microbial populations were quite different compared to
other samples. Brevibacillus brevis, Rhodospirillaceae bacterium, Bacillus spp, Vibrio spp and
Pseudomonas were present in all samples tested.

Chapter 7 PhyloChip
111
Chapter 7
Taxonomic analysis of Pentose-rich Natural Environments
Using a High Density Oligonucleotide Microarray
(PhyloChip) Technology
7.0 Summary
The microbial diversity of liquid samples was analysed in samples taken from the three main
sugar mills at Proserpine, Maryborough and Mackay in Queensland, Australia. Of particular
interest was the bagasse leachate sample, as sugar cane hemicellulose is the second most
dominant fraction of bagasse hemicellulose (23-30%), and it is mostly composed of xylose. The
main focus of this study was to explore the microbial diversity in a hemicelluloses-enriched
environment using the PhyloChip microarray identification system. A very complex microbial
community was found in each sample with the most divergent microbial community being found
in the bagasse leachate sample. The phylum Proteobacteria was the most abundant microbial
group present in all samples.
7.1 Introduction
The relatively recent introduction of molecular techniques for the detection and quantification
of microorganisms has started to permit a greater understanding of microbial diversity and its
role in nature. The most powerful new approach for the exploration of microbial diversity from
complex environmental samples is based on the cloning and sequencing of 16S ribosomal RNA
encoding genes. According to previous knowledge of the structure of these genes, coupled with
recent developments in PCR, it is now possible to identify closely related microorganisms by first
amplifying the 16S rRNA gene directly from isolated colonies using universal primers directed at
conserved regions at both ends of the gene, and then sequencing the PCR product (O'Sullivan,
2000).
Recently, a new method has been described, termed Comparative Genome Sequencing (CGS),
which resequences genomes based on custom oligonucleotide and cDNA hybridization arrays
generated by maskless photolithography, eliminating the need for constructing multiple physical
photolithography masks (Albert et al., 2005). More recently, the development of molecular
techniques, such as microarrays/metagenomic /environmental sequencing (Sessitsch et al.,

Chapter 7 PhyloChip
112
2006) or the new phylo sequencing methods, have shown potential advantages for bacterial
community characterization(Huse et al., 2008), and thus can be considered a new promising
direction for bacterial diversity monitoring. Indeed, a recent study showed that a microarray
targeting broad bacterial diversity could reveal greater diversity than had previously been
observed by the cloning-sequencing method (Loy et al., 2005). However, the metagenomic
PhyloChips developed so far focus mainly on functional bacterial groups or on single taxonomic
groups. Few microarrays have been developed in order to explore the microbial diversity (
Sanguin et al., 2009; Palmer et al., 2010). Diversity of communities can be described in terms of
taxa richness and taxa evenness. Richness provides an indication of total taxa present in a
sample/community, whereas evenness provides a measure of how dominant taxa are within a
community.
For the study reported in this chapter were analysed microbial communities in liquid samples
from sugar mills using a PhyloChip microarray analysis method which demonstrated the
presence of a possible pentose-capable community in the most potentially pentose-rich sample
(bagasse leachate).
7.2 Materials and Methods
7.2.1 DNA extraction from environmental samples and 16S rRNA gene amplification
Samples were collected from three different sugar mills at Proserpine (P), Mackay (MK) and
Maryborough (M) which are located in Queensland, Australia. Samples were: Floor dump sump
(FDS); cooling tower wash (CTW); and bagasse leachate (BGL). Seven liquid samples (sample
codes FDS-P, CTW-P, FDS-MK, CTW-MK, FDS-M, CTW-M and BGL-MK) were analysed by the
PhyloChip method. DNA was extracted from all liquid samples including bagasse leachate using
an ultra clean water DNA isolation kit (MOBIO laboratory inc. USA) and following the
manufacturer’s instructions. Extracted DNA was quantified by spectrophotometry to measure
the absorbance ratio at A260:A280, in order to determine the purity of the DNA.
Bacterial 16S rRNA genes were PCR amplified using primers 27F 5'(AGAGTTTGATCCTGGCTCAG)3'
and 1492R 5'(GGTTACCTTGTTACGACTT)3'. PCR mixtures included primers at 0.3 µM each,
dNTP’s at 200 μM each, 1.2 U of Taq polymerase (Takara), 10 x reaction buffers, 10 ng of
template DNA and water to 25 μL. Eight individual PCR reactions were set up over a primer
annealing range of 48-58°C (reducing primer bias). After hot-start enzyme activation, PCR
thermocycling consisted of 35 cycles of denaturation at 95°C for 30 s, annealing for 30 s and with
an extension at 72°C for 90 s. A final elongation step was performed for 10 min at 72°C. PCR

Chapter 7 PhyloChip
113
products from the separate 25 µL reactions were pooled, precipitated with isopropanol, washed
with 80% ethanol and resuspended in water to concentrate the PCR products for PhyloChip
analysis.
7.2.2 Microarray analysis of 16S rRNA genes
A high-density oligonucleotide microarray system (PhyloChip) was used to taxonomically identify
the 16S rRNA gene fragments from the liquid samples. The array system, built on the Affymetrix
GeneChip platform (supplied by Affymetrix CA, USA), has probes covering 8,741 prokaryotic 16S
rRNA OTU’s (Operational Taxonomic Unit) PhyloChip; (Brodie et al, 2006; Brodie et al., 2007 ).
For each sample, 500 ng of PCR product was mixed with a control oligonucleotide spike and
digested into 50-200 bp fragments with DNase I (Invitrogen) in One-Phor-All buffer (GE
HealthCare). The 3’ ends of the fragments were labelled with biotin using terminal
deoxynucleotidyl transferase (Promega) according to the GeneChip DNA labelling procedure
(Affymetrix, CA). The biotinylated mixture was denatured (99°C for 5 min) and then hybridised
to a PhyloChip microarray at 48°C and 60 rpm for 16 h. The hybridised array was washed and
stained (streptavidin-phycoerythrin) on an Affymetrix fluidics station according to protocols
described previously (Brodie et al., 2006 ).
7.2.3 Scoring taxa present
The raw PhyloChip array data (CEL data output files from the Affymetrix GeneChip Operating
System) were imported into PhyloTrac for analysis (chatz et al., 2010; PhyloTrac Environmental
Sample Analysis, 2011). Pixel images (fluorescence intensities in well defined grid) were resolved
as probe pairs (perfect match PM and mismatch MM), which are then grouped into probe sets
(OTUs). Each probe set contains an average of 24 probe-pairs per OTU and also contains a
central 17-mer not found in other OTUs. For each PM probe, a missmatched (MM) probe, with a
single nucleotide difference was present. For each probe set, the trimmed mean fluorescence
intensity (highest and lowest probe values removed before averaging) and normalized to
internal spike-in control intensities using a maximum likelihood method, then scaled to the
mean overall array and finally, log transformed. Taxa are considered present in a sample when
at least 90% of the probes in its probe set pass the following criteria: PM/MM ≥ 1.3 and PM ≥
130 x background noise(Desantis et al., 2007).
7.2.4 Classification of taxa
Based on probe sets found to be present on the array, PhyloTrac is immediately able to classify
bacterial taxa present in the samples, and also provide information on their taxonomic ranking

Chapter 7 PhyloChip
114
(i.e. phylum; class; order; family; sub-family data) (Schatz et al., 2010). This information was
exported into Excel in which manual reassignment of a few ambiguously-classified taxa was
conducted (e.g. some Bradyrhizobiaceae were classified into the Bradyrhizobiales order and
other Bradyrhizobiaceae into the Rhizobiales). The final tree, linking OTU intensity data in each
sample to taxonomic rankings, was imported into the Primer 6 software package (PRIMER-E Ltd,
UK), whereby data sets could be analysed at Phylum level through to sub-family level.
7.2.5 Diversity of bacteria present and identification of dominant taxa
The total number (i.e. taxa richness) of bacteria OTU’s and families were calculated in PhyloTrac.
The diversity, sensu community evenness/‘classical diversity’, was measured using the Shannon–
Weiner (Shannon; H’) index in Primer 6.
At each taxonomic level, the Weaver index was measured as:
Where pi is the proportion of intensity fluorescence for an individual OTU (at a given taxonomic
level) relative to the sum of all the intensities detected in the sample.
When determining which taxa of bacteria are dominant in a system, the overall intensity data
from PhyloChip cannot be directly used past OTU (sub-family) level. Aggregation of thosedata to
higher taxonomic levels imposes bias as probe coverage for bacteria such as Proteobacteria are
much greater than those for ‘rare’ groups such as Phylum TM7. This can be overcome by using
the ‘top 50’ taxa (probe intensity scores) for each ‘sample type’ (treatment). The most abundant
taxa are thereby selected and effects of differing coverage of probes across taxonomic groups
are minimized. Therefore, for each ‘sample type’ (treatment) the top 50 probe intensity values
were ranked and the combined data merged into a single file containing the top 50 for all
‘sample type’ (treatment) (total of 191 OTU). These data were used to explore which taxa were
dominant in abundance across the treatments. Taxonomic aggregation of OTUs was performed
in Primer6 as described previously.
7.2.6 Determining effects of treatment on bacterial community structure
The effects of treatment type on bacterial community assemblage were first explored at an OTU
level using 2-way clustering in PhyloTrac. Clustering of samples was based on Euclidean
distances and presented as a heat map.

Chapter 7 PhyloChip
115
In Primer6, individual data sheets were generated which contained PhyloChip OTU data of all
samples at each taxonomic level (i.e. data sheets at Phyla level, Order level etc), and the factors
‘sample type’ (treatment) and ‘location’ were assigned to the samples. For each data sheet, the
intensity values were log transformed and a resemblance matrix (similarity matrix) was
generated using the Bray-Curtis method. The influence of ‘sample type’ (treatment) and
‘location’ were then tested at each level of taxonomy using CAP analysis – canonical analysis of
principal components – with probability testing using permutation (999 repeats). Canonical
analysis of principle coordinates (CAP) using the software Primer 6.1 (http://www.primer-
e.com/). Data were log transformed prior to analysis and Euclidean distance was used to create
a similarity matrix comparing sample sites. Canonical analysis of principle were also carried out
as follows: relative taxon abundance was used as ‘species’ data, whereas liquid environmental
data were included in the analysis as ‘treatment’ variables. Correlations were considered
significant at a P<0.05 baseline and to be nearly significant at 0.05<P<0.10. Ordination methods
(nMDS, CAP) were then used to structure high-dimensional community composition data along
simple axes expressing overall compositional similarity and dissimilarity between sites.
Ordination via non-metric multidimensional scaling (nMDS) was used to the identify the effects
of ‘sample type’ (treatment) and ‘location’ detected following CAP analysis. Using PhyloChip
data aggregated to Class taxonomy, similarity between samples were calculated using the Bray-
Curtis method on Log-transformed intensity values. NMDS scaling was used to interpret the
distances in community composition (Bray-Curtis distances).
SIMPER analysis (similarity percentages / species contributions) was used to determine which
taxa were important in partitioning effects of sample treatment on bacterial community
composition (Clarke, 1993). All multivariate data analysis in the Primer6 software package
(PrimerE Ltd., UK) were conducted using routines described in (Clarke and Warwick, 2001).
7.3 Results
7.3.1 Richness and evenness of bacteria present
A DNA microarray (the G2 PhyloChip) was used as a tool to study the bacterial community
structure (Brodie, 2006). PhyloChip analysis revealed a high level of bacterial richness across all
samples as taxonomic groups. Richness of taxa was calculated based on scoring of PhyloChip
probe intensity data using PhyloTrac software package. Overall, 34 different Phyla were found to
be present (Table 7.1), which represents approximately half of those detectable by PhyloChip.
The total OTUs present was 1367 – approximately 16% of those covered on the array. The

Chapter 7 PhyloChip
116
highest richness of taxa was in the cooling tower water from Maryborough, which contained
nearly twice the OTUs than other samples, including the comparable sample from Mackay sugar
mill.
Table 7.1: Richness of the bacterial taxa present in environmental samples from sugar-cane
processing sites.
Level BGL-
MK
FDS-MK CTW-M CTW-MK FDS-M CTW-P FDS-P All
Phylum 29 30 32 21 30 21 13 34
Class 54 54 60 38 54 42 25 67
Order 106 90 117 58 90 62 32 127
Family 188 149 211 88 149 89 44 236
Sub-family 215 174 251 98 174 97 49 287
OTU 850 502 1023 209 502 218 135 1367
The microarray analysis also describes evenness of taxa distribution within communities
according to Shannon’s diversity index. Shannon’s index provides information on community
evenness as opposed to species richness. The higher the diversity indicated the higher
Shannon’s Index (Table 7.2). PhyloChip consistently detected more taxa in each of the samples
confirming the highest level of phylum and class found in cooling tower water sample from
Proserpine sugar mill (CTW-P). In addition, the highest number of order and family were found in
bagasse leachate from Mackay. The floor dump sump from Proserpine sugar mill showed the
highest variety of sub-families according to Shannon’s index. The PhyloChip-based OTU richness
according to Table 7.2 was 7.04±0.12. The communities are not being dominated by a few taxa
and this evenness is balanced across the samples.
Table 7.2: Evenness (Shannon’s diversity index; H’) of bacterial communities present from
sugar-cane processing sites.
Level BGL-
MK
FDS-MK CTW-M CTW-MK FDS-M CTW-P FDS-P
Phylum 1.64 1.83 1.71 1.67 1.77 1.85 1.68
Class 2.77 2.83 2.86 2.85 2.86 2.94 2.85
Order 3.94 3.70 3.89 3.98 3.84 3.80 4.01
Family 4.73 4.57 4.66 4.73 4.67 4.66 4.76
Sub-family 4.84 4.68 4.78 4.85 4.78 4.77 4.89
OTU 7.12 6.92 7.08 7.15 7.05 6.95 6.98

Chapter 7 PhyloChip
117
7.3.2 Identification of the dominant taxa
The top 60 PhyloChip probe intensity scores for each treatment were used to characterise the
dominant bacterial phyla in the samples. At a phylum level, the dominant bacteria presenting
Figure 7.1 were Firmicutes, Proteobacteria and Bacteroidetes.
However, the PhyloChip was able to detect a huge range of recessive phyla. Cyanobacteria,
Acidobacteria, TM7, Chlorobi, Natronoanaerobium, Spirochaetes and some unknown phyla were
the minor phyla, which had less than 50 PhyloChip intensity scores in all treatments.
Figure 7.1: Dominant bacterial phyla present. Samples collected from sugar-cane processing
sites, data based on combined top 50 probe intensity scores present in each sample.
BGL-MK

Chapter 7 PhyloChip
118
Figure 7.2: Distribution of bacterial Classes present within the four dominant Phyla.
From Figure 7.2 it is apparent that α-proteobacteria, β-proteobacteria, γproteobacteria, ϵ-
proteobacteria were present, which belong to the phylum Proteobacteria. Clostridium,
Bacillus, Mollicutes and Catabacter were in the Firmicutes groups. The only member of the
phylum Chloroflexi to be detected was the class Anaerolineae. There were three classes in
the Bacteroidetes groups: Bacteroidetes, Sphingobacteria and Flavobacteria.
The microarray detected probe intensity scores of 600,000 as shown in Figure 7.2,
Clostridium and Bacillus, which were the most dominant class in the floor dump sump
samples from the Mackay (FDS-MK) sugar mill. Bacillus, Clostridium, and α- and γ--
Proteobacteria were the most represented classes in the cooling tower waters from
Maryborough (CTW-M), and Mackay (CTW-MK) and also bagasse leachate from Mackay
sugar mills (BGL-MK). The same classes were present as dominant in the floor dump sump in
Maryborough (FDS-M), with the exception of β-Proteobacteria.
BGL-MK

Chapter 7 PhyloChip
119
The PhyloChip analysis revealed that Bacillus, Clostridium, and α-Proteobacteria were the
dominant classes in both the cooling tower water and the floor dump sump of the
Proserpine sugar mill. Exceptionally, there was some γ-proteobacteria found as a dominant
class in the floor dump sump from the Proserpine mill. The results presented in Figure 7.2
demonstrated that the most dominant phyla in all of the samples were Firmicutes and
Proteobacteria.
7.3.3 Determining effects of sample site on bacterial community structure
At the OTU level, the effects of the sampling sites were clearly distinguishable in terms of
community composition (Figure 7.3). For example, at this higher level of resolution the floor
dump sump in Maryborough had a distinctly different bacterial community from the other sites,
while the cooling tower water from Mackay mill exhibited high overall diversity at a taxonomic
level.

Chapter 7 PhyloChip
120
Figure 7.3: Heat map / 2-way clustering of samples according to PhyloChip OTU’s. Clustering
based on Euclidean distance using complete linkage method.
The influence of treatment and location were then formally tested using CAP analysis. CAP
analysis was performed at all taxonomic levels and the summary effects data are presented in
Table 7.3. Since there was a single sample for bagasse leachate, CAP analysis was only
performed to explore differences between the floor dump sump and cooling tower water
treatments and across each of the three locations (Mackay, Maryborough and Prosperine).

Chapter 7 PhyloChip
121
Table 7.3: Summary of CAP data showing the significance of sample types and location effects
on bacterial community composition
Taxa level P ( samples type )(treatment) P (location)
Phylum 0.196 0.795
Class 0.012 0.809
Order 0.087 0.861
Family 0.127 0.640
Sub-family 0.281 0.505
Highlighted cell is a highly significant effect
CAP analysis showed that at very low taxonomic level (sub-family, family) there were no main
treatment effects. The treatment effect means the effect of sample collection site could be
found. There was no significant difference of the diversity of taxa level on the basis of the variety
of the sample types (floor dump sump, cooling tower water and bagasse leachete). This is
surprising as, at this level, most families will be present across the samples. In the middle levels
(Class level) of the taxonomic heirarchy, the effects of sample type (but not location) were highly
significant. At the highest level (Phylum) this relationship is lost, probably as there are not
enough potential differences between Phyla for variations to be observed.
Accordingly, at the Class-level of taxonomy only, ordination of bacterial community structure by
non-metric multidimensional scaling (nMDS) was conducted.

Chapter 7 PhyloChip
122
treatment
Floor dump sediment
Cooling tower
Bagasse
Maryborough
Maryborough
Mackey
Proserpine
ProserpineMaryborough
Mackey
2D Stress: 0.001
Mackay
Mackay
Mackay
Maryborough
Proserpine
Proserpine
Maryborough
treatment
Floor dump sediment
Cooling tower
Bagasse
Maryborough
Maryborough
Mackey
Proserpine
ProserpineMaryborough
Mackey
2D Stress: 0.001
Figure 7.4: nMDS ordination plot showing similarity in bacterial community structure (Class
level) between samples. Samples in close proximity have a more similar community composition.
The nMDS ordination demonstrated the effect (left to right) of treatments on bacterial
community structure (Figure 7.4). A change in taxa composition can be seen in samples moving
from cooling tower water, to floor dump sump, to bagasse leachate. Microbial diversity in
bagasse samples is more divergent due to different habitat composition.
Based on the finding of a highly significant treatment effect on the distribution of bacterial
Classes between cooling tower water and floor dump sump samples, SIMPER analysis was used
to determine which Classes contributed most strongly to the treatment effects. Of the 67
classes detected, variation in only about seven of these contributed to 20% of the total
‘treatment’ effect variation. The summary SIMPER data are given in Table 7.4.

Chapter 7 PhyloChip
123
Table 7.4: Summary of SIMPER data. Table allocates differences of bacterial classes towards
describing the overall ‘treatment’ or ‘sampling site' effect between floor dump and cooling
tower samples on bacterial community composition.
Floor dump sump Cooling tower water
Classes Av. Abund Av .abund Contrib% Cum.%
NC10-1 8.81 10.19 4.94 4.94
Acidobacteria-4 10.28 10.93 3.32 8.26
Betaproteobacteria 14.62 15.38 3.05 11.31
Proteobacteria (UC) 11.55 12.01 2.62 13.93
Deinococcus-Thermus (UC) 11.53 12.20 2.40 16.33
Flavobacteria 13.73 14.34 2.27 18.60
TM7 (UC) 9.99 10.43 2.12 20.71
UC=Un-cultured class
Av.abund = average abundance
Contribut% = percent contribution of that Class to the “sample type” effect
Cum% = cumulative percentage of Classes
The data show that differences in abundance of bacterial Class NC10-1 are most important
(contributing to about 5%) in describing the overall differences in bacterial community
composition between the treatments, however the magnitude of even this effect was small
(only ~5%). Cumulative variation in the abundance of a number of different bacterial classes was
needed to describe the treatment effect more fully. Interestingly, these classes were from a
number of different Phyla.
NC10-1 is merely a candidate Class of the NC10 phyla (there are 2 classes NC10-1 and NC10-2).
None of these exists in pure culture. They are only known by DNA-based detection.
7.4 Discussion
From the earliest cultivation experiments to today's metagenomic analyses, most of the major
discoveries in this field were driven by applications of novel methods. Molecular ecology has had
a major impact by revealing the true scope of microbial diversity and providing genetic markers
that could be used to track important species, even in cases where cultures were unavailable
(Marco, 2010). A phylogenetic microarray that has been developed to used as an example of a
microarray that targets the known diversity within the 16S rDNA gene to determine microbial
community composition (Andersson et al., 2008). Using multiple, confirmatory probes to

Chapter 7 PhyloChip
124
increase the confidence of detection and a mismatch probe for every perfect match probe to
minimize the effect of cross-hybridization by non-target regions.
Bacterial community analysis demonstrated the diversity of bacterial DNA in a hemicelluloses-
enriched environment. PhyloChip microarray data demonstrated a very complex bacterial
ecosystem (details discussed in next paragraph) present in the environment related to the
Queensland sugar industry, in particular the mills. Only seven liquid samples including bagasse
leachate were analysed by using the PhyloChip microarray method. Half of these samples were
identified by the DGGE and 16S rDNA sequencing method; and rest of these samples were
analysed by using PhyloChip microarray. PhyloChip mostly defines bacteria to the phylum, order
and sometimes class level. Bacterial species present in seven liquid samples, including bagasse
leachate, were described in chapter 6 (Table 6.3) as determined by 16S rDNA sequencing.
Acidobacteria, Betaproteobacteria, Deinococcus thermus and Flavobacteria were mostly found
in these selected samples.
According to the PhyloChip analysis, the Phyla Proteobacteria and Firmicutes were the most
dominant groups present in all samples. The Proteobacteria phylum includes a wide range of
pathogens, such as Escherichia coli, Salmonella sp., Vibrio sp., Helicobacter sp. etc. (Madigan
and Martinko, 2005). However, the majority of species in this Phylum are free-living, and include
a number of nitrogen fixing bacteria. According to the information from DGGE and 16S rDNA
sequencing, Vibrio sp. was common species in all liquid samples. Rhodospirillaceae sp. and
Azospirillum brasilense are also a part of the Proteobacteria that were found present in samples
by 16S rDNA sequence analyses.
Proteobacteria comprise one of the largest Phyla of prokaryotes and account for the vast
majority of the known (culturable) Gram-negative bacteria. This group of organisms
encompasses a very complex assemblage of phenotypic and physiological attributes including
many phototrophs, heterotrophs and chemolithotrophs. The Phylum Proteobacteria is of great
biological significance as it includes a large number of known human, animal and plant
pathogens.
The Phylum Firmicutes includes all of the Gram-positive bacteria, but has recently been
redefined to include a core group of related forms called the low-G+C group, in contrast to the
Actinobacteria (Haakensen et al., 2008). Many Firmicutes produce endospores, which are
resistant to desiccation and can survive extreme conditions, making them ideal for
environmental ecosystems such as soil and water. Members of the Firmicutes are known to be

Chapter 7 PhyloChip
125
principally responsible the degradation of major polysaccharides forming acetate, formate, lactic
acid etc (Teusink and Smid, 2006). The presence of Firmicutes as a dominant group is quite rare
for natural samples. In the sugar mill environment, the dominance of Firmicutes may be linked
to the presence of ‘lactic acid’ type bacterial communities. Lactobacilli commonly share the
habitat when the content of sugar level is high in that habitat (Kleerebezem and Vaughan, 2009).
It has been mentioned earlier that bagasse leachate contains very high levels of hemi-cellulose.
According to Kleerebezem and Vaughan (2009), lactic acid bacteria are beneficial Firmicutes in
the intestinal tract, which produce lactic acid as an end-product of carbohydrate fermentation.
This ability has been exploited in food production owing to the growth-inhibiting effect that
acidification has on spoilage agents. Since the lactic acid bacteria are generally regarded as safe
(GRAS) for specifically defined uses and they often produce narrow spectrum antibacterial
peptides active against pathogenic bacteria, they have also been used for health-promoting
purposes such as probiotics (Teusink et al, 1998; Teusink and Smid, 2006). Thus the presence of
high levels of Firmicutes in sugar mill waste samples may have significant implications in the
nutriceuticals industry.
Firmicutes occupy a wide variety of habitats, and can be either useful or problematic in various
food and beverage related industries (Sakamoto and Konings, 2003), in the fuel alcohol industry
(Skinner and Leathers, 2004), and in human and animal health (Carr et al., 2002). It is believed
that numerous industrial applications of Firmicutes such as lactic acid production, remain to be
exploited (Teusink et al., 1998; Teusink and Smid, 2006). One prominent Firmicute, Bacillus spp.,
was found in all samples as a very common bacterium in accord with the results from Chapter 6.
Burkholderia cepacia is a member of the sub-phylum β-proteobacteria and was found in the
floor dump sump of Mackay mill (FDS-MK). Alcaligenes faecalis, also a member of same phylum,
was found in bagasse leachate only. However, species belonging to the phylum, Acidobacteria,
such as Deinococcus thermus and Flavobacteria were not found by 16S rDNA sequencing. This
might be due to experimental error either with DGGE or 16S rDNA sequencing (chapter 6) or
simply that these were present in numbers too low to detect. So this finding reveals that, the
PhyloChip analysis is able to explore wider microbial diversities compared to DGGE methods.
In this study, the Phyla Proteobacteria and Firmicutes were present as the most dominant
groups in bagasse. These include the sub-groups α-proteobacteria, β-proteobacteria, Clostridium
and Bacillius. Interestingly, Firmicutes are numerically low dominant groups in bagasse leachate
sample compared to other samples.This is an interesting finding. The bagasse leachate sample is
the most different in terms of community composition. The subsequent analysis could only look

Chapter 7 PhyloChip
126
for changes between the floor dump sump (FDS) and cooling tower water (CTW) samples which,
on the nMDS are a little bit different but not nearly as much as the bagasse leachate. This may
be because of the composition of nutrients in the habitat. Alcaligenes faecalis was found only in
bagasse leachate. This information revealed that baggasse leachate might be enriched in carbon
and nitrogen sources (hemicellulose).
Lignocellulosic materials, such as sugar cane bagasse represent an abundant, inexpensive source
of organic material that can be a carbon source for microbial growth. Sugarcane bagasse
leachate is residual liquid in nature, created by rainwater passing through the bagasse stockpile
and contains dissolved lignocelluloses. Sugar cane bagasse wastes are an additional source of
microbial nutrients (nitrogen and phosphate source). The micro flora may have the ability to
grow in lignocellulose type substrates. The spent compost/bagasse mix, besides being an
important source of nitrogen and phosphate nutrients, also provides an appropriate support for
the solid matrix as well as native microorganisms that are capable of pentose-utilization.
The class level of taxonomy was highly significant according to the analysis of sample type. The
nature of the floor dump sump (FDS) and cooling tower water (CTW) were different. Basically
microorganism found in the floor dump sump is most likely to be composed of soil bacteria and
that of cooling tower water is most likely composed of airborne bacteria. This may be the
possible reason why there is a high significance of microbial diversity at the class level in these
two types of samples. It was also demonstrated by MDS analysis (Figure 7.4), that the bagasse
leachate from Mackay mill was the most different but it was not possible to compare the nature
of this, because only one sample was available, and from just one location.
Cooling tower water from Maryborough contained the highest richness of taxa according to the
richness analysis. Microorganisms enter cooling towers through the water supply. The constant
fall of water within cooling towers make an air scrubber and which delivers large amounts of
organic, inorganic particulates and microorganisms into the water phase. The combination
increases the water temperatures, humidity and large surface areas in cooling towers to provide
an ideal environment for microbial growth.
7.5 Conclusions
All of the samples studied by the PhyloChip microarray system were highly rich in taxa, which
included not only the numerically dominant groups, but also rarer taxa, indicative of a very
complex bacterial ecosystem. The bagasse leachate sample was the most different in terms of
community composition.

Chapter 7 PhyloChip
127
The richness analysis represents that the cooling tower water from Maryborough contained the
highest richness of taxa compared to other samples, which was unexpected. According to the
evenness analysis (Shannon’s Index), the highest level of phylum and class were found in cooling
tower water from Proserpine (CTW-P) and the highest number of order and family were found in
bagasse leachate from Mackay (BGL-MK). In addition, the highest variety of sub-families was
found in the floor dump sump of Proserpine mill. Firmicutes, Proteobacteria and Bacteroidetes
were present as a dominant phylum in all samples according to the analysis of the identification
of the dominant taxa (Figure 7.1). Moreover, CAP analysis found no significance difference of the
diversity of taxa level on the basis of variety of the sampling location. At the class level of
taxonomy, the effects of sample type (but not location) were highly significant (P=0.12).
The PhyloChip is able to simultaneously identify many thousands of taxa present in an
environmental sample. The PhyloChip is shown to reveal greater diversity within a community
than rRNA gene sequencing due to the placement of the entire gene product on the microarray
compared with the analysis of up to thousands of individual molecules by traditional sequencing
methods (Liu and Jansson, 2010).

Chapter 8 Summary
128
Chapter 8
Summary, conclusion and future work
8.1 Revisiting the hypothesis and aims
The research presented in this thesis was conducted with the aim of investigation of the
hypothesis which was: “That pentose-utilizing organisms may be isolated from pentose-rich
natural environments and consequently be used to ferment pentose sugars such as those
found in hemicellulose from agricultural waste”. It was further hypothesized that the pentose
degradation process would result in commercially valuable end-products such as amino acids.
This hypothesis was investigated by addressing three major aims. The first aim of this research
was to isolate, identify and characterize Gram-positive microbes which were able to degrade
pentose sugars as a carbon source in the culture medium.
The second aim of this research was to detect and identify amino acids as a major end-product
from single pentose cultivation and dual sugar (pentose plus glucose) cultivation of the E-PUB
isolates obtained. The diauxie characteristics and specific growth rates from single and dual
carbon sources in the growth medium were also investigated.
The final aim was to investigate the diversity of microbial populations in pentose-enriched
habitats, using metagenomic methods such as DGGE and PhyloChip microarrays. The dominant
microbial species were identified using 16S rDNA sequencing methods.
8.1.1 Hypothesis
The hypothesis was proved. Six Gram-positive microbes, which belong to the Order
Actinomycetales, were isolated from pentose-rich agricultural waste material. They were able to
utilize pentose sugars at approximately the same rate and efficiency as they utilize glucose. The
extension of the hypothesis was also proved: pentose-utilizing microbes were able to produce
commercially valuable amino acids, in this case glycine.

Chapter 8 Summary
129
8.2 Summary of findings
Pentose-degrading microbes were isolated from the pentose-enriched environment. Six
indigenous Gram-positive microbes were isolated, and were found to belong to the order
Actinomycetales but were of various genera and species. These microbes were isolated from the
soil of sugar treatment ponds from the Proserpine and Maryborough sugar mills. The six
isolated microbes were screened from 191 isolates on the basis of their physiological and
biochemical characteristics. Finally these six indigenous bacteria were identified to the species
level, using 16S rDNA sequencing analysis.
It was confirmed that the isolated microbes were Corynebacterium glutamicum,
Corynebacterium freiburgense, Actinomyces odontolyticus, Rhodococcus equi, Nocardia elegans,
and Propionibacterium freudenreichii. These are of interest as they are uncommon, but
important species in the pentose-enriched environment, as shown by ecological metagenomic
studies.
It was also investigated whether the isolated microbes were able to grow with pentose sugars as
a carbon source in a minimal culture medium, to simulate the growth conditions in their native
habitat. It was proven that these natural isolates were able to use pentose as a carbon source
during growth.
The focus of further study was on the simulated utilization of lignocellulosic biomass through
cultivation. Of the three major components of such biomass, cellulose has almost all carbon in
the form of glucose, whereas hemicellulose also has some pentose. The medium composition
was created in order to simulate the sugar content of “natural” sugarcane bagasse, which
consists of 25.2% xylose and 41.0% glucose, expressed as % w/w of the dry matter (Pandey et
al., 2000).
Consequently, the pentose sugars (e.g., xylose, arabinose, and ribose) nearly always constitute a
much smaller proportion of lignocellulosic biomass than do hexoses. Moreover, microorganisms
generally prefer to metabolise hexoses over pentoses. However, our findings show that the
isolates C. glutamicum, C. freiburgense, A. odontolyticus, R. equi, N.elegans, P. freudenreichii
were able to metabolise pentose approximately same efficiency rate as they did glucoseutilizing
pentose sugars (xylose, arabinose or ribose) as a single sole carbon source during growth
without any genetic modification. All environmental pentose-utilizing bacteria (E-PUB) were able
to metabolise xylose efficiently, as shown by the specific growth rate, which was approximately
same for pentose sugars as for glucose metabolism.

Chapter 8 Summary
130
E-PUB microorganisms were grown on pentoses or hexoses as single carbon and energy sources.
Growth with glucose resulted in a slightly higher specific growth rate than that for xylose or
arabinose.
Dual sugar utilization was also tested. It was proven that all E-PUB microorganisms were able to
show diauxie characteristics during growth on dual sugar combinations (glucose plus
xylose/arabinose/ribose). Diauxie growth with similar growth rate was observed when glucose
was mixed with either xylose or arabinose or ribose and the glucose was consumed first. The
beginning of consumption of the second substrate generally followed a lag phase that was
longer than the lag phase for the first substrate, after which consumption of the two sugars
proceeded simultaneously. Our research suggests that mixtures of hexoses and pentoses, such
as those present in hemicellulose-hydrolysates, can be broken down by the natural E-PUB
microorganisms, thus proving that they are good candidates for the biodegradation of
hemicellulose-hydrolysates. Further research into improving pentose consumption is indicated,
as well as extending the complexity of the carbon source mixture to one approaching that found
in hemicellulose-hydrolysates.
Following the tests that demonstrated the ability of natural isolates to use pentose sugars in the
presence and absence of glucose, it was investigated the end-products of such metabolism. The
amino acid end-products were detected and analysed using High Performance Liquid
Chromatography (HPLC). It was shown that all six isolated microbes were able to produce
arginine and cysteine after growth on xylose, except for N. elegans which did not produce any
cysteine. C. glutamicum and P. freudenreichii produced glycine after single pentose metabolism
under oxygen deprivation conditions. P. freudenreichii also produced alanine after a single sugar
was consumed. An anoxic condition following an aerobic culture period allows the metabolism
of the substrate with the resulting production of industrially valuable amino acids. All six
isolates produced glycine after dual (xylose and glucose) sugar catabolism. P. freudenreichii was
able to degrade arabinose and also produce glycine after dual sugar (arabinose plus glucose)
utilization. Glycine has considerable importance in pharmaceutical applications. Arginine and
cysteine both have importance in food and pharmaceutical industries.
The microbial biosynthesis pathway of glycine, cysteine, arginine and alanine (Stanier et al.,
1977) shows that glycine, and alanine are amino acids of the serine and pyruvate family. The
formation of these amino acids is carried out via the pentose phosphate pathway. Thus, finding
this group of amino acids as end-products in the fermentative process is not unexpected,
particularly using the pentose phosphate pathway for the metabolism of pentose sugars. The

Chapter 8 Summary
131
relationship between the various amino acids and the final product glycine also indicates a
shared biosynthetic pathway.
An ecological study was performed using Denaturing Gradient Gel Electrophoresis (DGGE), and
16S rDNA sequencing methods. This provided further insight into the bacterial community
composition of the solid and liquid samples from the sugar mills, and the similarities and
differences of the genotypic patterns for each of the bacterial communities. The objective of this
study was to evaluate the species diversity contained in a complex habitat. These metagenomic
methods are useful to investigate poorly described, biochemically deficient, or fastidious
organisms. The results of sequencing are more accurate due to the database of phylogenetic
relationships available (Mignard and Flandrois, 2006).
The Phylum Proteobacteria was the most common microbial phylum found in this experiment.
The Actinomycetes group was not dominant in the eco-niches tested using DGGE, despite their
culturability in early experiments. It would appear that the more abundant groups of micro-
organisms in both soil and water samples did not respond to the culture conditions provided,
either because of sub-lethal damage due to drought conditions, or because the culture
conditions did not suit their requirements. Thus, even though the six E-PUB microorganisms
belonged to the family Actinomycetes, this was not a dominant microbial group in the sites from
which these samples were selected. However, Corynebacterium sp., which is related or similar to
the six E-PUB isolates, was also detected in mud and raw bagasse samples. These six E-PUB
microorganisms were the end result of screening 191 microorganisms found in sugar mill
environmental sources (pond soil). Most of the species of Corynebacterium, Nocardia and
Rhodococcus are widely distributed in the environment, but particularly abundant in soil (Holt,
2000).
The significance of the relative rarity of the E-PUB species is that, in order to assess their
capacity in raw material (in particular, bagasse), the raw material will need to be seeded with
the chosen organisms for use in a commercial fermentation process. Due to the relatively small
population of these organisms, a natural seeding process would not be efficient enough to
permit the industrial processing of bagasse. The diversity of microbial communities was also
investigated using High Density Oligonucleotide Microarray (PhyloChip) Technology. The
information about the size of the microbial diversity in liquid samples from the sugar mills was
significant. Acidobacteria, Firmicutes, Beta-Proteobacteria, Deinococcus thermus and
Flavobacteria groups were found in most of the selected samples.

Chapter 8 Summary
132
Microorganisms belonging to the actinomycetes group were not found by PhyloChip assay. This
could be due to the relatively low abundance of the actinomycetes group in the samples tested
as this was also demonstrated to be the case using DGGE analysis. It also revealed that the
microbial diversity depends on the nutritional composition of the natural habitat.
Corynebacterium spp were commonly found in culture based identification, DGGE and PhyloChip
method from sugar mills due to the similarity of habitat. The organisms found in the bagasse
leachate, the most likely pentose-rich sample tested, were significantly different to those of the
floor dump sump and the cooling tower water. However, very common environmental and
highly rich taxa were demonstrated by using this microarray system.
8.3 Significance of findings
This project has made significant contributions to the recycling of agricultural waste material, by
examining its usage as a substrate for industrial fermentation processes, particularly with
respect to the fraction of hemicellulose-, and the pentose sugars, which are abundant in this
source.
Six indigenous bacteria were isolated and identified from the environment, and were able to use
pentose sugars without any genetic modification. They were able to utilize pentose in the
presence of glucose, as would be found in their native environment, and also in its absence. The
efficiencies of the utilization of pentose (xylose, arabinose, and ribose) and glucose were about
equal. Most importantly, the fermentative process resulted in a valuable commercial product,
namely the amino acid glycine. Quantitative analysis also has completed, there was high
concentrationof amino acids produced by E-PUB.
Furthermore, the ecology of the micro-community present in environmental samples from
pentose-rich habitats was investigated using metagenomic methods, and it was established that
they were very complex. The indigenous pentose-utilizing bacteria that were isolated (E-PUB)
were not dominant in the eco-systems tested, with the exception of Corynebacterium sp, which
has implications for the industrial outcomes of this research.
To conclude, the E-PUB isolated in this research were able to utilize pentose naturally. While it
has been found previously that many microorganisms can utilize pentose after genetic
modification, this is the first report of organisms of the Corynebacterium group that are able to
do this without genetic manipulation. While genetically modified organisms can have many
advantages in industrial processes, one major disadvantage is that they may back-mutate to
form a wild type organism during the rather long fermentation period required for this particular

Chapter 8 Summary
133
process (Anderson and Moselio, 2009). Thus, the use of natural, non-mutated organisms is a
great advantage.
8.4 Future directions
Industrial fermentation will be the main outcome of this research. Future work may focus on
determining the relative size of populations of these different bacteria in order to understand
the possibility of a dynamic relationship, illustrated by their presence in leachate and bulk
material and their presence and numbers at specific stages of the degradation process.
To achieve the highest growth rate on pentose-containing feed-stock, and higher product yield,
extensive trials will need to be undertaken. These will determine the impact of changes of
temperature, pH, nutritional composition and incubation period on biomass formation and feed-
stock degradation. In addition, enzymatic analysis should be undertaken to understand the
activity of enzymes responsible for the degradation of pentoses and examine the possibility of
up-regulating them. Finally, quantitative analysis of substrate sugars consumed and end product
analysis also will be a part of the future work.
To maximise the use of biomass, one to ten percent inoculum of microbial cells is generally used
as a starter, to begin the initial aerated step. The optimum growth conditions for C. glutamicum,
N. elegans, P. freudenreichii are: temperature of 30°C; pH 6, in the presence of oxygen (Haynes
and Britz, 1990; Zhao et al., 2011; Lemee et al., 1994). Actinomyes odontolyticus is a newly
identified bacterium. The optimal growth conditions of this bacterium are: temperature range
35-37°C;pH 6-7; and facultative anaerobic condition (Ramos et al., 1997). The growth conditions
need to be optimized for the industrial fermentation using E-PUB. Such conditions as pH;
availability of suitable levels of nutrients such as nitrogen, phosphorus and trace elements;
optimum temperature range, oxygen availability during growth and depletion during
fermentation all must be optimized. In particular, the pH should be at the optimum level during
growth, but during end-product production the pH may be reduced (e.g. citric acid production
from Aspergillus niger; (Haq et al., 2003; Berovic et al., 2007; Sikander et al., 2002). It was not
possible to complete the optimization of the many growth and fermentation variables, due to
time constraints, however it is obvious that this is an important future research objective.
Future experiments should include determination of the efficiency of utilization of different
carbon sources in complex sources such as molasses and other agricultural by-products. For
example Brevibacterium sp. utilises different carbon sources (glucose, fructose, sucrose,
maltose, lactose, xylose and starch) for its growth and for the production of glutamic acid. It is

Chapter 8 Summary
134
possible that our E-PUB organisms may be capable of utilizing a wider range of carbon sources,
which would permit a wider application of the intended industrial process.
Bagasse leachate can be used as a substrate, however before fermentation starts, the bagasse
may need to be treated to destroy the unwanted wild microorganisms present, although ideally
the E-PUB organisms will function effectively in a “natural” substrate. If using heat to treat the
bagasse leachate, the sugars present will be polymerized, and the carbon could be rendered
unsuitable for use by microorganisms. Steam fractionation could be an alternative. Timing is a
main issue for steam fractionation. Steam fractionation of bagasse leachate could be an
expensive process for the scaling up of end-product formation. According to Sasaki et al. (2003),
bagasse samples were extracted as a water-soluble or steam fraction at low temperatures (200–
230°C), and about 30% was extracted at higher temperatures (230–280°C). At 200–230°C,
hydrolysates of hemicellulose (galactose, arabinose and xylose) and aromatic compounds mainly
existed in the extract solution. At this stage, hardly any glucose and cellobiose, which are
normally formed via hydrolysis of cellulose, were yielded and few aromatic compounds were
formed.
Another step would be the measurement of cfu/mL during biomass production. Industrial
fermentation uses fed-batch cultivation of microbes or other single cells, occurring firstly with
and then without, the presence of air. The aeration is performed by stirring the culture mix, or
bubbling air through it, but often using both methods simultaneously. The levels of stirring and
bubbling would need to be optimized for each process. Fed-batch fermentation is an industrial
fermentation technique in between batch and continuous fermentation (Hong, 1986; Stanbury
et al., 1993). A controlled feed rate, with the right component constitution is required during the
process. Fed-batch offers many advantages over batch and continuous cultures. From the
concept of its implementation it can be easily concluded that under controlled conditions and
with the required knowledge of the microorganism involved in the fermentation, the feed of the
required components for growth and/or other substrates required for the production of the
product can never be depleted and the nutritional environment can be maintained
approximately constant during the course of the batch. Sometimes, controlling the substrate is
also important due to catabolite repression. Since this method usually permits the extension of
the operating time, high cell concentrations can be achieved and thereby, improved productivity
(mass of product/volume x time). This aspect is greatly favored in the production of growth-
associated products (Stanbury et al., 1993).
Furthermore, in a fed-batch fermentation, no specialized equipment is required in addition to
that required for batch fermentation, even considering the operating procedures for sterilization

Chapter 8 Summary
135
and the preventing of contamination (Longobardi, 1994). A cyclic fed-batch culture has an
additional advantage: the productive phase of a process may be extended under controlled
conditions. The controlled periodic shifts in growth rate provide an opportunity to optimize
product synthesis, particularly if the product of interest is a secondary metabolite whose
maximum production takes place during the deceleration in growth (Stanbury et al., 1993).
Following the creation of biomass, the process is changed to an anoxic state by stopping the
bubbler to allow microbes to produce fermentation products. The shaking should continue, to
prevent the biomass from settling to the floor of the vessel, which is known as a precipitation.
The process is not unlike that of beer production, in which, following aerobic biomass
production, the yeast is stirred slightly in order to remain fully distributed through the beer
during the anaerobic fermentation step (McPhee, 2003).
Intracellular amino acids were analyzed in this research. As discussed above, different carbon
sources as growth supplements were used so that the possibility of different end-products could
be explored. Examples of bacteria that have proved useful in this context previously include:
Nocardia sp that have industrial importance for the production of antibiotics (Aggarwal et al.,
2011; Chiba et al., 2007); P. freudenreichii, which is important to produce acetic acid and
propionic acid (Lemee et al., 1994); and Corynebacterium glutamicum that is widely used for the
industrial production of amino acids and nucleotides (Haynes and Britz, 1990). Intracellular end-
products were extracted in this study though microorganisms are able to produce both
intracellular and extracellular products. Intracellular end-product analysis could be a significant
contribution in future research as Nocardia, Corynebacterium and Propionibacterium are known
to be able to produce intracellular amino acids (Wittmann et al., 2004; Limpisathian, 2005). C.
glutamicum, Propionibacterium sp are also able to produce extracellular amino acids (Wittmann
et al., 2004; Limpisathian, 2005). In addition, Actinomyces spp. produce cysteine as an
intracellular and valine as an extracellular by products (Eriquez and Pisano, 1979; Schaal et al.,
2006).
In the future, it will be possible to focus on the production of other organic acid and amino acid
end-products after the fermentation of complex carbon sources found in agricultural waste
materials. A diagram of the entire project, including both the completed work and future
directions is shown in Figure 8.1.

Chapter 8 Summary
136
Figure 8.1: Project outlook

References
137
References
Agbogbo, F. K., Kelly-Coward, G., Torry-Smith, M. and Wenger, K. S. 2006. "Fermentation of
glucose/xylose mixtures using pichia stipitis". Process Biochemistry 41: 2333-2336.
Aggarwal, K., Jing, F., Maranga, L. and Liu, J. 2011. "Bioprocess optimization for cell culture based
influenza vaccine production". Vaccine In Press, Corrected Proof.
Albert, T. J., Dailidiene, D., Dailide, G., Norton, J. E., Kalia, A., Richmond, T. A., Molla, M., Singh, J.,
Green, R. D. and Berg, D. E. 2005. "Mutation discovery in bacterial genomes: Metronidazole
resistance in helicobacter pylori". Nature Methods 2 (12): 951-953.
Almersheim, P. 1976. "The primary cell wall. In: "Plant biochemistry"". Ed. Bonner, J., Varner, J. E.
(Eds.) 3rd. ed. ed. New York, London: Academic Press.
Amann, R. I., Ludwig, W., Schleifer, K. H. 1995. "Phylogenetic identification and in situ detection of
individual microbial cells without cultivation". Microbiology and Molecular Biology Reviews
59 (1): 143-169.
Amore, R., Kotter, P., Kuster, C., Ciriacy, M. and Hollenberg, C. P. 1991. "Cloning and expression
in Saccharomyces cerevisiae of the NAD(P)H-dependent xylose reductase-encoding gene
(XYL1) from the xylose-assimilating yeast Pichia stipitis". Gene 109: 89–97.
Andersson, A.F., Lindberg, M., Jakobsson, H., Backhed, F., Nyren, P., Engstrand, L. 2008.
"Comparative analysis of human gut microbiota by barcoded pyrosequencing”. PLoS ONE
3:e2836
Anderson, T. M. and Moselio, S. 2009. "Industrial fermentation processes". In Encyclopedia of
microbiology, 349-361. Oxford: Academic Press.
Atlas, M. R. 2004. Handbook of microbiological media: CRC press.
Attfield, P. V. and Philip, J. L. B. 2006. "Use of population genetics to derive nonrecombinant
saccharomyces cerevisiae strains that grow using xylose as a sole carbon source". Federation
of European Microbiological Societies: 862–868.
Aubel, D., Germond, J.E., Gilbert, C., and Atlan, D. 2002. "Isolation of the patC gene encoding
the cystathionine beta-lyase of Lactobacillus delbrueckii subsp. bulgaricus and molecular
analysis of inter-strain variability in enzyme biosynthesis”. Microbiology 148(7):2029-
2036.
Barker, T. W. and Worgan, J. T. 1981. "The utilisation of palm oil processing effluents as substrates
for microbial protein production by the fungus aspergillus oryzae". Applied Microbiology and
Biotechnology 11 (4): 234-240.
Beaman, B.L., Kim, S.K., Salton, M.R.J., and Barksdale, L. 1971. "Amino acids of the cell wall of
Nocardia rubra “. The Journal of Bacteriology 108: 941–943.

References
138
Beaman, B.L. 1975. "Structural and biochemical alterations of nocardia asteroids cell walls during its
growth cycle". The Journal of Bacteriology: 1235-1253.
Becker, J. and Boles, E. 2003. "A modified saccharomyces cerevisiae strain that consumes l-arabinose
and produces ethanol". Applied Environmental Microbiology 69: 4144 - 4150.
Belitz, H. D., Grosch, W. and Schieberle, P. 2004. "Food chemistry". Springer.
Berovic, M. L., Matic; M. Raafat El-Gewely. 2007. "Citric acid production". Biotechnology annual
review, 303-343: Elsevier.
Barry, D.P., Beaman, B.L. 2006. “Modulation of eukaryotic cell apoptosis by members of the bacterial
order Actinomycetales”. Apoptosis 11(10):1695-707.
Billard P., Menart, S., Fleer, R . and Bolotin-Fukuhara, M. 1995." Isolation and characterization of the
gene encoding xylose reductase from Kluyveromyces lactis" Gene 162: 932e .
Bosshard, P. P., Abels, S., Zbinden, R., Böttger, E. C. and Altwegg, M. 2003. "Ribosomal DNA
sequencing for identification of aerobic gram positive rods in the clinical laboratory (an 18-
months evaluation)". Journal of Clinical Microbiology 41: 4134–4140.
Bosshard, P. P., Abels, S., Altwegg, M., Böttger, E.C., Zbinden, R. 2004. "Comparison of conventional
and molecular methods for identification of aerobic catalase negative gram positive cocci in
the clinical laboratory". Journal of Clinical Microbiology 42: 2065–2073.
Bothast, R. J., Nichols, N. N. and Dien, B. S. 1999. "Fermentations with new recombinant organisms".
Biotechnology Progress 15: 867 - 875.
Brodie, E. L., Desantis, T.Z., Joyner, D.C., Baek, S.M., Larsen, J.T., Andersen, G.L., Hazen, T.C.,
Richardson, P.M., Et Al. 2006. "Application of a high-density oligonucleotide microarray
approach to study bacterial population dynamics during uranium reduction and
reoxidation". Applied Environmental Microbiology 72: 6288-6298.
Brodie, E. L., Desantis, T.Z., Parker, J.P.M., Zubietta, I.X., Piceno, Y.M., Andersen, G.L. 2007. "Urban
aerosols harbor diverse and dynamic bacterial populations". Ed. Sci104, P. N. A. USA.
Brons, J. K., and Van Elsas, J.D. 2008. "Analysis of bacterial communities in soil by use of denaturing
gradient gel electrophoresis and clone libraries, as influenced by different reverse primers".
Applied Environmental Microbiology 74: 2717-2727.
Brückner, R., and Titgemeye, F. 2002. "Carbon catabolite repression in bacteria: Choice of the carbon
source and autoregulatory limitation of sugar utilization.” FEMS Microbiology Letter 209:
141-148.
Bruinenberg, P. M., Bot, P. H. M., Dijken, J. P. and Scheffers, W. A. 1983. "The role of redox balances
in the anaerobic fermentation of xylose by yeasts". Applied Microbiology and Biotechnology
18 (5): 287-292.
Brownc, M. and Rosea, . H. 1969b. "Fatty acid composition of Candida utilis as affected by
growth temperature and dissolved oxygen tension". Journal of Bacteriology 97: 371-378.
Buckeridge, K. M. and Grogan, P. 2008. "Deepened snow alters soil microbial nutrient limitation
in arctic birch hummock tundra". Appllied Soil Ecology. 39: 210–222.

References
139
Burkovski, A., (Editor) 2008. "Corynebacteria: Genomics and molecular biology." Caister Academic
Press.
Carr, F. J., Chill, D. And Maida, N. 2002. "The lactic acid bacteria: A literature survey." Critical Reviews
in Microbiology 28: 281–370.
Castro, D. F. H., Oliveira, C. S., and Furlan, A. S. 2003. "Alternative approach for utilization of pentose
stream from sugarcane bagasse by an induced flocculent Pichia stipitis". Applied
Biochemistry and Biotechnology 105 (108): 547-555.
Chaabane, F. B., Aldiguier. S. A., Alfenore, S., Cameleyre, X., Blanc, P., Bideaux, C., Guillouet, E. S.,
Roux, G., Molina-Jouve, C. 2006. "Very high ethanol productivity in an innovative continuous
two-stage bioreactor with cell recycles." Bioprocess and Biosystems Engineering 29 (1).
Chandel, A.K., Singh, O.V., Chandrasekhar, G., Rao, L.V., Narasu, M.L. 2010c. “Bioconversion of
novel substrate, Saccharum spontaneum, a weedy material into ethanol by Pichia stipitis
NCIM3498”. Biores Technol (in press)
Chiba K, H. Y., Ishino K, Kogure T, Mikami Y, Uehara Y, Ishikawa J 2007. "Construction of a pair of
practical Nocardia-Escherichia coli shuttle vectors". Japanese Journal of Infectious Disease 60
(1): 45–47.
Christon, J. H. 1991. "Modeling the environmental fate of microorganisms." Washington, DC:
Published American Society for Microbiology.
Clarke, K. R. 1993. "Non-parametric multivariate analysis of changes in community structure".
Australian Journal of Ecology 18: 117–143.
Clarke, K. R., Warwick, R.M. 2001. "Change in marine communities: An approach to statistical
analysis and interpretation". Plymouth, UK: second ed Primer-E Ltd.
Clarridge, J. E. 2004 "Impact of 16s rRNA gene sequence analysis for identification of bacteria on
clinical microbiology and infectious diseases". Clinical Microbiology Reviews 17: 840-862.
Colwell, R.R., Spira, W.M., and D. Barua. 1992. The ecology of Vibrio cholerae. In Current Topics in
Infectious Disease: Cholera. Eds Barua and Greenaugh
Connon, S. A., Tovanabootr, A., Dolan, M., Vergin, K., Giovannoni, S. J. and Semprini, L. 2005.
"Bacterial community composition determined by culture-independent and -dependent
methods during propane-stimulated bioremediation in trichloroethene-contaminated
groundwater". Environmental Microbiology 7 (2): 165-178.
Craig, J. W., Chang, F.-Y., Kim, J. H., Obiajulu, S. C. and Brady, S. F. 2010. "Expanding small-molecule
functional metagenomics through parallel screening of broad-host-range cosmid
environmental DNA libraries in diverse proteobacteria". Applied Environmental Microbiology
76 (5): 1633-1641.
CSR Ethanol Facts. 2001. "Part 5 - Sugar Milling Process"
http://www.ethanolfacts.com.au/sugarprocess. ( Accessed February 22nd, 2011).
Cybercolloids. "Introduction to pectin - structure".

References
140
http://www.cybercolloids.net/library/pectin/introduction-pectin-structure (Accessed February 24th,
2011).
Daley, P., Petrich, A., May, K., Luinstra, K., Rutherford, C., Chedore, P., Jamieson, F. and Smieja, M.
2008. "Comparison of in-house and commercial 16S rRNA sequencing with high-
performance liquid chromatography and genotype AS and CM for identification of
nontuberculous Mycobacteria". Diagnostic Microbiology and Infectious Disease 61 (3): 284-
293.
Daßler, T., Maier, T., Winterhalter, C. and Böck, A. 2000. "Identification of a major facilitator protein
from Escherichia coli involved in efflux of metabolites of the cysteine pathway". Molecular
Microbiology 36 (5): 1101-1112.
De La Cruz-Canizares, J. Gimeno-Adelantado, J.V., Mateo-Castro, R., Bosch-Reig, F., Domenech-
Carbo, M.T. 2004. "Suppression of pigment interference in the gas chromatographic analysis
of proteinaceous binding media in paintings with EDTA". Journal of Chromatography A 1025
( 2): 277-285.
Dees, P. M. and Ghiorse, W. C. 2001. "Microbial diversity in hot synthetic compost as revealed by
PCR-amplified rRNA sequences from cultivated isolates and extracted DNA". FEMS
Microbiology Ecology 35 (2): 207-216.
Demain, A. L. 2005. "Microbial cells and enzymes: a century of progress". Methods in Biotechnology
17: 1-27.
Demoling, F. 2007. "Comparison of factors limiting bacterial growth in diff erent soils". Soil
Biology Biochemistry 39: 2485–2495.
Desantis, T., Brodie, E., Moberg, J., Zubieta, I., Piceno, Y. and Andersen, G. 2007. "High-density
universal 16S Rrna microarray analysis reveals broader diversity than typical clone library
when sampling the environment". Microbial Ecology 53 (3): 371-383.
Dhillon, S. G., Nagasawa, T. and Yamada H. 1987. "Microbial process for L-cysteine production".
Enzyme and Microbial Technology. 9(5):277-280.
Dien, B.S., Nichols, N., O'Bryan, P. and Bothast, R. 2000. "Development of new ethanologenic
Escherichia coli strains for fermentation of lignocellulosic biomass". Applied Biochemistry
and Biotechnology 84-86 (1): 181-196.
Dien, B.S., Cotta, M. A. and Jeffries, T. W. 2003. "Bacteria engineered for fuel ethanol production:
Current status". Applied Microbiology and Biotechnol ogy 63: 258 - 266.
Dien, B. S., Nichols, N. N. and Bothast, R. J. 2002. "Fermentation of sugar mixtures using Escherichia
coli catabolite repression mutants engineered for production of l -lactic acid". Journal of
Industrial Microbiology and Biotechnology 29 (5): 221-227.
Di Luccio, E., Elling, R. A. and Wilson, D. K. 2006. "Identifi cation of a novel NADH-specifi c aldo-
keto reductase using sequence and structural homologies". Biochemistry Journal
400:105–114.

References
141
Embley, T. M. and Stackebrandt, E. 1994. "The molecular phylogency and systematics of the
Actinomycetes". Annual Review of Microbiology 48 (1): 257-289.
Ercolini, D. 2004. "PCR-DGGE fingerprinting: Novel strategies for detection of microbes in food".
Journal of Microbiological Methods 56 (3): 297-314.
Eriquez, L. A. and Pisano, M.A. 1979. "Isolation and nature of intracellular alpha-aminoadipic acid-
containing peptides from paecilomyces persicinus P-10". Antimicrobial Agents and
Chemotherapy: 392-397.
Euzéby, J. P. 2011. "List of Prokaryote names with standing in Nomenclature"
http://www.bacterio.cict.fr/number.html#total. (Accessed February, 2011).
Fong, P. 2008. "Hungry for answers: Wheat Kings Reign in time of need" Toronto Star, published
Monday, July 7, 2008. . Accessed October 2010. http://www.thestar.com/news/canada/
article/455433--wheat-kings-reign-in-time-of-need.
Frank, M. P. and Powers, R. W. 2007. "Simple and rapid quantitative high-performance liquid
chromatographic analysis of plasma amino acids". Journal of Chromatography B 852 (1-2):
646-649.
Freeborn, R. A., West, K. A., Bhupathiraju, V. K., Chauhan, S., Rahm, B. G., Richardson, R. E. and
Alvarez-Cohen, L. 2005. "Phylogenetic analysis of tce-dechlorinating consortia enriched on a
variety of electron donors". Environmental Science & Technology 39 (21): 8358-8368.
Funke, G., Frodl, R., Bernard, K. A. and Englert, R. 2009. "Corynebacterium freiburgense sp. Nov.,
isolated from a wound obtained from a dog bite". International Journal of Systematic and
Evolutionary Microbiology 59 (8): 2054-2057.
Gatti, R. and Gioia, M. G. 2008. "Liquid chromatographic fluorescence determination of amino acids
in plasma and urine after derivatization with phanquinone". Biomedical Chromatography 22
(2): 207-213.
Gatti, R., Gioia, M. G., Leoni, A. and Andreani, A. 2010. "2,5-dimethyl-1h-pyrrole-3,4-dicarbaldehyde
as a precolumn derivatization reagent for HPLC/UV detection of amino acids". Journal of
Pharmaceutical and Biomedical Analysis 53 (2): 207-211.
Ghrist, C. A. and George, V. Stauffer.1995." The Escherichia coli glycine transport system and its
role in the regulation of the glycine cleavage enzyme system" Microbiology 141:133-140.
Gírio, F. M., Fonseca, C., Carvalheiro, F., Duarte, L. C., Marques, S. and Bogel-Lukasik, R. 2010.
"Hemicellulose for fuel ethanol: A review". Bioresource Technology 101 (13): 4775-4800.
Glickman, S. E., Kilburn, J. O., Butler, W. R. and Ramos, L. S. 1994. "Rapid identification of mycolic
acid patterns of mycobacteria by high-performance liquid chromatography using pattern
recognition software and a mycobacterium library". Journal of Clinical Microbiology 32 (3):
740-745.
Goodfellow, M., Alderson, G. and Chun, J. 1998. "Rhodococcal systematics: Problems and
developments". Antonie van Leeuwenhoek 74 (1): 3-20.

References
142
Govindaswamy, S. and Vane, L. M. 2007. "Kinetics of growth and ethanol production on different
carbon substrates using genetically engineered xylose-fermenting yeast". Bioresource
Technology 98 (3): 677-685.
Green, S. J., Michel, F. C., Jr., Hadar, Y. and Minz, D. 2007. "Contrasting patterns of seed and root
colonization by bacteria from the genus Chryseobacterium and from the family
Oxalobacteraceae". International Society for Microbial Ecology Journal 1 (4): 291-299.
Grimmler, C., Held, C., Liebl, W. and Ehrenreich, A. 2010. "Transcriptional analysis of catabolite
repression in Clostridium acetobutylicum growing on mixtures of d-glucose and d-xylose".
Journal of Biotechnology 150 (3): 315-323.
Gundersen, R. Y., Vaagenes, P., Breivik, T., Fonnum, F. and Opstad, P. K. 2005. "Glycine – an
important neurotransmitter and cytoprotective agent". Acta Anaesthesiologica Scandinavica
49 (8): 1108-1116.
Haakensen, M., Dobson, C. M., Deneer, H. and Ziola, B. 2008. "Real-time PCR detection of bacteria
belonging to the firmicutes phylum". International Journal of Food Microbiology 125 (3):
236-241.
Hahn-Hägerdal, B., Galbe, M., Gorwa-Grauslund, M. F., Lidén, G. and Zacchi, G. 2006. "Bio-ethanol -
the fuel of tomorrow from the residues of today". Trends in Biotechnology 24 (12): 549-556.
Hahn-Hägerdal, B., Karhumaa, K., Fonseca, C., Spencer-Martins, I. and Gorwa-Grauslund, M. 2007.
"Towards industrial pentose-fermenting yeast strains". Applied Microbiology and
Biotechnology 74 (5): 937-953.
Hahn-Hagerdal, B., Wahlbom, C. F., Gardonyi, M., Van Zyl, W. H., Cordero Otero, R. R. and Jonsson, L.
J. 2001. "Metabolic engineering of Saccharomyces cerevisiae for xylose utilization". Advances
in Biochemical Engineering Biotechnology 73: 53 - 84.
Handumrongkul, C., Ma, D. P. and Silva, J. L. 1998. "Cloning and expression of Candida
guilliermondii xylose reductase gene (xyl1) in Pichia pastoris." Applied Microbiology and
Biotechnology 49:399-404.
Haq, I.-U., Ali, S. and Iqbal, J. 2003. "Direct production of citric acid from raw starch by Aspergillus
niger". Process Biochemistry 38 (6): 921-924.
Harvey, L. M. and McNeil, B. 1998. "Thickeners of microbial origin". In Microbiology of fermented
foods, ed. B., W. J. London: Blackie Academic & Professional.
Haynes, J.A, and Britz, M. 1990. "The effect of growth conditions of Corynebacterium glutamicum on
thetransformation frequency obtained by electroporation". Journal of General Microbiology
136: 255-263.
Heinze, T., Ebringerová, A. and Hromádková, Z. 2005. "Hemicellulose". In Polysaccharides i, 1-67:
Springer Berlin / Heidelberg.
Held, P. 2012. “Enzymatic Digestion of Polysaccharides, Part II: Optimization of Polymer
Digestion and Glucose Production in Microplates”. Biofuel Research.
Henderson, W. 2009. "Agilent application Note, 5990-4547en ".

References
143
Herberdt, D. 1961. "The chemical composition of micro-organisms as a function of their
environment". Symposia of the Society for General Microbiology II 391-416.
Hermann, T. 2003. "Industrial production of amino acids by coryneform bacteria". Journal of
Biotechnology 104 (1-3): 155-172.
Hoch, G. 2007. "Cell wall hemicellulose as mobile carbon stores in non-reproductive plant tissues".
Functional Ecology 21 (5): 823-834.
Holt, J. G. ed. 2000. Bergey’s Manual of Determinative Bacteriology.
Hong, J. 1986. "Optimal substrate feeding policy for a fed batch fermentation with substrate and
product inhibition kinetics". Biotechnology and Bioengineering 28 (9): 1421-1431.
Hornsey, I. S. 2003. " A history of beer and brewing". UK : Royal Society of Chemistry: Cambridge.
Horst-Backhaus, B. E., Felske, A., Snaidr, J., Wieshuber, A., Amann, R., Ludwig, W., and Nu¨ Bel, U.
1996. "Sequence heterogeneities of genes encoding 16S rRNAs in Paenibacillus polymyxa
detected by temperature gradient gel electrophoresis". Journal of Bacteriology 178 (19):
5636–5643.
Hui, Y.H., Nip, W.K., Rogers, R., and Young, O.A. (Eds). 2001. Meat Science and Applications. New
York: Marcel Dekker.
Hundal, H.S. and Taylor, P.M. 2009. "Amino acid transceptors: Gatekeepers of nutrient exchange
and regulators of nutrient signaling". American Journal of Physiology – Endocrinology &
Metabolism 296: E603-E613.
Huse, S. M., Dethlefsen, L., Huber, J. A., Welch, D. M., Relman, D. A. and Sogin, M. L. 2008. "Exploring
microbial diversity and taxonomy using SSU rRNA hypervariable tag sequencing". PLoS
Genetics 4 (11): e1000255.
Huyghe, A., Francois, P., Charbonnier, Y., Tangomo-Bento, M., Bonetti, E.-J., Paster, B. J., Bolivar, I.,
Baratti-Mayer, D., Pittet, D., Schrenzel, J., et al. 2008. "Novel microarray design strategy to
study complex bacterial communities". Applied Environmental Microbiology 74 (6): 1876-
1885.
Ikeda, M. and Katsumata, R. 1999. "Hyperproduction of tryptophan by Corynebacterium glutamicum
with the modified pentose phosphate pathway". Applied Environmental Microbiology 65 (6):
2497-2502.
Ingram, L. O., Conway, T., Clark, D. P., Sewell, G. W. and Preston, J. F. 1987. "Genetic engineering of
ethanol production in Escherichia coli". Applied Environmental Microbiology 53: 2420 - 2425.
Inui, M., S., Murakami, S., Okino, H., Kawaguchi, A. A., Vertès, and H. Yukawa. 2004. "Metabolic
analysis of Corynebacterium glutamicum during lactate and succinate productions under
oxygen deprivation conditions". Journal Molecular Microbiology and Biotechnology 7: 182-
196.
Iwatani S, Dien VS, Shimbob K, Kubota K, Kageyamab N, Iwahata D, Miyanob H, Hirayama K,
Usuda Y , ShimizuK , Matsui K. (2007). "Determination of metabolic flux changes during
fed-batch cultivation from measurements of intracellular amino acids by LC-MS/MS".

References
144
Journal of Biotechnology 128: 93–111.
Janssen, L. and Moscicki, L. 2009. "Hermoplastic starch: A green material for various industries".
Netherlands: Wiley-VCH.
Janssen, M., Slenders, P., Tramper, J., Mur, R. L. and Wijffels, H. René. 2001. "Photosynthetic
efficiency of Dunaliella tertiolecta under short light/dark cycles". Enzyme and Microbial
Technology 29 : 298-305.
Jeffries, T. W., Grigoriev, I. V., Grimwood, J., Laplaza, J. M., Aerts, A., Salamov, A., Schmutz, J.,
Lindquist, E., Dehal, P., Shapiro, H., et al. 2007. "Genome sequence of the lignocellulose-
bioconverting and xylose-fermenting yeast Pichia stipitis". Nature Biotechnology 25 (3): 319-
326.
Jeppsson, M., Bengtsson, O., Franke, K., Lee, H., Hahn-Hagerdal, B. and Gorwa-Grauslund, M. F.
2006. "The expression of a Pichia stipitis xylose reductase mutant with higher k(m) for
NADPH increases ethanol production from xylose in recombinant Saccharomyces cerevisiae".
Biotechnology and Bioengineering 93: 665-673.
Jeppsson. M., Träff, K., Johansson, B., Hahn-Hägerdal, B., Gorwa-Grauslund, M.F. 2003."Effect of
enhanced xylose reductase activity on xylose consumption and product distribution in
xylose-fermenting recombinant Saccharomyces cerevisiae". FEMS Yeast Ressearch 3:167-
175.
Jez, J. M., Bennett, M. J., Schlegel, B. P., Lewis, M., and Penning, T. M. 1997. "Comparative
anatomy of the aldo-keto reductase superfamily" Biochemistry Journal 326: 625.
John, R., Nampoothiri, K. and Pandey, A. 2007. "Fermentative production of lactic acid from biomass:
An overview on process developments and future perspectives". Applied Microbiology and
Biotechnology 74 (3): 524-534.
Johnston, S. "Queensland's sugar industry". Source: http://www.johnston-independent.com/
sugar.html (Accessed February 12th, 2008).
Jump, D. B., Clarke, S. D., Thelen, A., and Liimata. M. 1994. "Coordinate regulation of glycolytic and
lipogenic gene expression by polyunsaturated fatty acids". Journal of. Lipid Reseasrch.
35:1076-1084.
Kalinowski, J., Bathe, B., Bartels, D., Bischoff, N., Bott, M., Burkovski, A., Dusch, N., Eggeling, L.,
Eikmanns, B. J., Gaigalat, L., et al. 2003. "The complete Corynebacterium glutamicum ATCC
13032 genome sequence and its impact on the production of -aspartate-derived amino acids
and vitamins". Journal of Biotechnology 104 (1-3): 5-25.
Kanekar, P. P., Nilegaonkar, S. S., Sarnaik, S. S. and Kelkar, A. S. 2002. "Optimization of protease
activity of alkaliphilic bacteria isolated from an alkaline lake in india". Bioresource
Technology 85 (1): 87-93.
Karhumaa, K., Hahn-Hagerdal, B. and Gorwa-Grauslund, M. F. 2005. "Investigation of limiting
metabolic steps in the utilization of xylose by recombinant Saccharomyces cerevisiae using
metabolic engineering". Yeast 22: 359 - 368.

References
145
Kastner, J. R., Jones, W. J. and Roberts, R. S. 1999. "Ethanol fermentation of mixed-sugars using a
two-phase, fed-batch process: Method to minimize d-glucose repression of Candida
shehatae d-xylose fermentations". Journal of Industrial Microbiology and Biotechnology 22
(2): 65-70.
Kawaguchi, H., Vertes, A. A., Okino, S., Inui, M. and Yukawa, H. 2006. "Engineering of a xylose
metabolic pathway in Corynebacterium glutamicum". Applied Environmental Microbiology
72 (5): 3418-3428.
Kim, J.W., Kim, H.J., Kim, Y., Lee, M.S., Lee, H.S. 2001."Properties of the Corynebacterium
glutamicum metC gene encoding cystathionine beta-lyase".Molecular Cells 11(2):220-
225.
Kim, Y., Ingram, L. O. and Shanmugam, K. T. 2007. "Construction of an Escherichia coli k-12 mutant
for homoethanologenic fermentation of glucose or xylose without foreign genes". Applied
Environmental Microbiology 73 (6): 1766-1771.
Kirchman LD. and Hodson ER. 1986. "Metabolic regulation of amino acid uptake in marine
waters". Limnol Oceanogr 31(2): 339-350
Kirchner, O. and Tauch, A. 2003. "Tools for genetic engineering in the amino acid-producing
bacterium Corynebacterium glutamicum". Journal of Biotechnology 104 (1-3): 287-299.
Kleerebezem, M., & Vaughan, E.E. 2009 Probiotic and gut lactobacilli and bifidobacteria: Molecular
approaches to study diversity and activity. Ann Rev Microbiol 63: 269–290
Kratzer, R., Kavanagh, K. L., Wilson, D. K. and Nidetzky, B. 2004. "Studies of the enzymic
mechanism of Candida tenuis xylose reductase (AKR 2B5): X-ray structure and catalytic
reaction profile le for the H113A mutant". Biochemistry 43:4944–4954.
Koukkou, A. ed. 2011. Microbial bioremediation of non-metals: Caister Academic Press.
Kuan, Y.J., Charnley, S.B., Huang, H.C., Tseng, W.L., and Kisiel, Z. 2003. "Interstellar glycine ". The
Astrophysical Journal 593 (2): 848-867.
Lacey, J. I97I, "Thermoactinomyces sacchari sp.nov. a thermophilic actinomycete causing
bagassosis". Journal of General Microbiology 66,327-338.
Laser, M., Schulman, D., Allen, S. G., Lichwa, J., Antal, M. J. and Lynd, L. R. 2002. "A comparison of
liquid hot water and steam pretreatments of sugar cane bagasse for bioconversion to
ethanol". Bioresource Technology 81 (1): 33-44.
Laurent, F. J., Provost, F. and Boiron, P. 1999. "Rapid identification of clinically relevant Nocardia
species to genus level by 16S rRNA gene pcr". Journal of Clinical Microbiology 37 (1): 99-102.
Lebo, S. E. Jr.; Gargulak, J. D. and McNally, T. J. 2001. "Lignin". In Kirk-Othmer Encyclopedia of
Chemical Technology. John Wiley & Sons, Inc.
Lee, J. 1997. "Biological conversion of lignocellulosic biomass to ethanol". Journal of Biotechnology
56 (1): 1-24.

References
146
Lefebvre, D., Gabriel, V., Vayssier, Y. and Fontagné-Faucher, C. 2002. "Simultaneous HPLC
determination of sugars, organic acids and ethanol in sourdough process". Lebensmittel-
Wissenschaft und-Technologie 35 (5): 407-414.
Lemee, R., Lortal, S., Cesselin, B., and van Heijenoort. J. 1994. "Involvement of an n-
acetylglucosaminidase in autolysis of Propionibacterium freudenreichii cnrz 725". Applied
and Environmental Microbiology 60 (12): 4351-4358.
Leuchtenberger, W., Huthmacher, K. and Drauz, K. 2005. "Biotechnological production of amino
acids and derivatives: Current status and prospects". Applied Microbiology and
Biotechnology 69 (1): 1-8.
Li, Z., Xu, J., Tang, C., Wu, J., Muhammad, A. and Wang, H. 2006. "Application of 16S rDNA-PCR
amplification and DGGE fingerprinting for detection of shift in microbial community diversity
in Cu-, Zn-, and Cd-contaminated paddy soils". Chemosphere 62 (8): 1374-1380.
Liang, Z., Drijber, R. A., Lee, D. J., Dwiekat, I. M., Harris, S. D. and Wedin, D. A. 2008. "A DGGE-cloning
method to characterize arbuscular mycorrhizal community structure in soil". Soil Biology and
Biochemistry 40 (4): 956-966.
Ligthelm, M. E., Prior, B. A. and Preez, J. C. 1988. "The oxygen requirements of yeasts for the
fermentation of d-xylose and d-glucose to ethanol". Applied Microbiology and Biotechnology
28 (1): 63-68.
Liles, M. R., Turkmen, O., Manske, B. F., Zhang, M., Rouillard, J.-M., George, I., Balser, T., Billor, N.
and Goodman, R. M. 2010. "A phylogenetic microarray targeting 16S rRNA genes from the
bacterial division Acidobacteria reveals a lineage-specific distribution in a soil clay fraction".
Soil Biology and Biochemistry 42 (5): 739-747.
Limpisathian, P. 2005. Characterization of the interaction between Lactobacillus helveticus and
Propionibacterium in Swiss cheese. Doctor of Philosophy, Food Science and Nutrition
Graduate Program, Ohio State University.
Lin, Y. and Tanaka, S. 2006. "Ethanol fermentation from biomass resources: Current state and
prospects". Applied Microbiology and Biotechnology 69 (6): 627-642.
Liu, J., Barry, C. E., Besra, G. S. and Nikaido, H. 1996. "Mycolic acid structure determines the fluidity
of the mycobacterial cell wall". Journal of Biological Chemistry 271 (47): 29545-29551.
Liu, W.-T. and Jansson, J. K. 2010. "Environmental molecular microbiology". Berkeley: Caister
Academic Press.
Longobardi, P. G. 1994. "Fed-batch versus batch fermentation. " Bioprocess Engineering 10: 185 -
194.
Loy, A., Schulz, C., Lucker, S., Schopfer-Wendels, A., Stoecker, K., Baranyi, C., Lehner, A. and Wagner,
M. 2005. "16S rRNA gene-based oligonucleotide microarray for environmental monitoring of
the betaproteobacterial order "Rhodocyclales". Applied Environmental Microbiology 71 (3):
1373-1386.
Lu, X., Sun, J., Nimtz, M., Wissing, J., Zeng, A.P., Rinas, U. 2010. "The intra- and extracellular
proteome of Aspergillus niger growing on defined medium with xylose or maltose as

References
147
carbon substrate". Microbial Cell Factories 9(23): 1-13.
Lynd, L.R., L., K., South, C.R., Van, Walsum, G.P., Levenson, K. 2001. "Evaluation of paper sludges for
amenability to enztmatic hydrolysis and conversion to ethanol". Tippi’s Journal 84 (2): 50.
Madigan, M. and Martinko, J. eds. 2005. Brock's biology of microorganisms. Prentice-Hall.
Marco, D. ed. 2010. Metagenomics: Theory, methods and applications. Argentina: Caister Academic
Press.
Mallory, L.M., Austin, B., and R.R. Colwell. 1977. "Numerical taxonomy and ecology of oligotrophic
bacteria isolated from the estuarine environment". Canadian Journal of Microbiology, 23(6): 733-
750
Mateos, L. M., Ordóñez, E., Letek, M. and Gil, J. A. 2006. "Corynebacterium glutamicum as a model
bacterium for the bioremediation of arsenic". International Microbiology 9 (3).
McCowen, M. S. and Phibbs, V. P., JR. 1974."Regulation of alanine dehydrogenase in Bacillus
licheniformis". Journal of Bacteriology 118(2):590-597.
McLeod, M. P., Warren, R. L., Hsiao, W. W. L., Araki, N., Myhre, M., Fernandes, C., Miyazawa, D.,
Wong, W., Lillquist, A. L., Wang, D., et al. 2006. "The complete genome of Rhodococcus sp.
Rha1 provides insights into a catabolic powerhouse". Proceedings of the National Academy
of Sciences 103 (42): 15582-15587.
McPhee, J. 2003. "The unacknowledged workers of the biotechnology industry: Microbes." In The
Science Creative Quarterly, August 2003 edition. British Columbia, Canada.
Meinhard,S.,1991."Isotope dilution of intracellular amino acids as a tracer of carbon and
nitrogen sources of marine planktonic bacteria". Marine Ecology Progress Series 74:
295-301.
Mignard, S. and Flandrois, J. P. 2006. "16S rRNA sequencing in routine bacterial identification: A 30-
month experiment". Journal of Microbiological Methods 67 (3): 574-581.
Mohagheghi, A., Evans, K., Chou, Y.-C. and Zhang, M. 2002. "Co-fermentation of glucose, xylose, and
arabinose by genomic DNA-integrated xylose/arabinose fermenting strain of Zymomonas
mobilis ax101". Applied Biochemistry and Biotechnology 98-100 (1): 885-898.
Moore, S. and Stein, W.H. 1951. "Chromatography of amino acids on sulfonated polystyrene resins ".
The Journal of Biological Chemistry 192 (663).
Morbach, S. and Krämer, R. 2003. "Impact of transport processes in the osmotic response of
Corynebacterium glutamicum". Journal of Biotechnology A New Era in Corynebacterium
Glutamicum Biotechnology 104 (1-3): 69-75.
Mordarski, M., Goodfellow, M., Tkacz, A., Pulverer, G. and Schaal, K. P. 1980. "Ribosomal ribonucleic
acid similarities in the classification of Rhodococcus and related taxa". Journal of General
Microbiology 118 (2): 313-319.
Morimoto, S., Togami, K., Ogawa, N., Hasebe, A. and Fujii, T. 2005. "Analysis of a bacterial
community in 3-chlorobenzoate-contaminated soil by PCR-DGGE targeting the 16S rRNA

References
148
gene and benzoate 1,2-dioxygenase gene (bena)". Microbes and Environments 20 (3): 151-
159.
Motoyama, H., Maki, K., Anazawa, H., Ishino, S., Teshiba, S. 1994. "Cloning and nucleotide
sequence of the Homoserine dehydrogenase genes (hom) and the threonine synthase
genes (thrC) of the gram-negative obligate methylotroph Methylobacillus glycogens".
Applied Environmental Microbiology 60: 111-119.
Muhling, M., Woolven-Allen, J., Murrell, J. C. and Joint, I. 2008. "Improved group-specific PCR
primers for denaturing gradient gel electrophoresis analysis of the genetic diversity of
complex microbial communities". International Society for Microbial Ecology Journal 2 (4):
379-392.
Muyzer, G. and Smalla, K. 1998. "Application of denaturing gradient gel electrophoresis (DGGE) and
temperature gradient gel electrophoresis (TGGE) in microbial ecology". Antonie van
Leeuwenhoek 73 (1): 127-141.
My Agriculture Information Bank/Soil Microbiology
( http://agriinfo.in/?page=topic&superid=5&topicid=147) Accessed 21/07/2012.
Nampoothiri, K. M. and Pandey, A. 1995. "Effect of different carbon sources on growth and glutamic
acid fermentation by Brevibacterium sp." Journal of Basic Microbiology 35 (4): 249-254.
Naono, S., Rouviere, J. and Gros, F. 1965. "Preferential transcription of the lactose operon during the
diauxic growth of Escherichia coli". Biochemical and Biophysical Research Communications
18 (5-6): 664-674.
Nathan-Mosier, C. W., Dale, B., Elander, R., Lee, Y., Holtzapple, M. and Ladisch, M. 2005. "Features of
promising technologies for pretreatment of lignocellulosic biomass." Bioresource Technology
96 (6).
Neufeld, J. D., Mohn, W. W. and De Lorenzo, V. 2006. "Composition of microbial communities in
hexachlorocyclohexane (hch) contaminated soils from spain revealed with a habitat-specific
microarray". Environmental Microbiology 8 (1): 126-140.
Neihardft, C. 1963. "Effect of environment on the composition of bacterial cells". Annual Review
of Microbiology 17: 61-86.
NH&MRC. 2004. "Australian drinking water guidelines 6". Trans. Council, A. G. N. R. M. M., National
Water Quality Management Strategy.
National Renewable Energy Laboratory (NREL). 1999. "Bioethanol multi-year technical plan: Fiscal
year 2000 and beyond". Golden, Colorado.
Nishizawa, M., Katou, Y., Shirahige, K. and Toh-e, A. 2004. "Yeast Pho85 kinase is required for proper
gene expression during the diauxic shift". Yeast 21 (11): 903-918.
Nunan, N., Daniell, T. J., Singh, B. K., Papert, A., McNicol, J. W. and Prosser, J. I. 2005. "Links between
plant and rhizoplane bacterial communities in grassland soils, characterized using molecular
techniques". Applied Environmental Microbiology 71 (11): 6784-6792.

References
149
Nunn, B.L. and Keil, R. G. 2006. "A comparison of non-hydrolytic methods for extracting amino acids
and proteins from coastal marine sediments." Marine Chemistry 98 (1): 31-42.
O'Sullivan, D. J. 2000. "Methods for analysis of the intestinal microflora". Current Issues in Intestinal
Microbiology 1 (2): 39–50.
Öhgren, K., Bengtsson, O., Gorwa-Grauslund, M. F., Galbe, M., Hahn-Hägerdal, B. and Zacchi, G.
2006. "Simultaneous saccharification and co-fermentation of glucose and xylose in steam-
pretreated corn stover at high fiber content with Saccharomyces cerevisiae tmb3400".
Journal of Biotechnology 126 (4): 488-498.
Ohnishi, J., Katahira, R., Mitsuhashi, S., Kakita, S. and Ikeda, M. 2005. "A novel gnd mutation leading
to increased l-lysine production in Corynebacterium glutamicum". FEMS Microbiology Letters
242 (2): 265-274.
Okabe, S. and Kamagata, Y. 2010. "Wastewater treatment". In Environmental molecular
microbiology, eds. Liu, W.-T. and Jansson, J. K.: Caister Academic Press.
Olsson, L. and Hahn-Hägerdal, B. 1996. "Fermentation of lignocellulosic hydrolysates for ethanol
production". Enzyme and Microbial Technology 18 (5): 312-331.
Ozcan, S., P. Kotter, and M. Ciriacy. 1991. " Xylan-hydrolyzing enzymes of the yeast Pichia stipitis".
Applied Microbiology and Biotechnology 36: 190-195.
Palmer, C., Bik, E. M., Eisen, M. B., Eckburg, P. B., Sana, T. R., Wolber, P. K., Relman, D. A. and Brown,
P. O. 2010. "Rapid quantitative profiling of complex microbial populations". Nucleic Acids
Research 34 (1): e5.
Pandey, A., Soccol, C. R., Nigam, P. and Soccol, V. T. 2000. "Biotechnological potential of agro-
industrial residues. I: Sugarcane bagasse". Bioresource Technology 74 (1): 69-80.
Patel, J. B. 2001. "16S rRNA gene sequencing for bacterial pathogen identification in the clinical
laboratory". Molecular Diagnosis 6 (4): 313-321.
Patel, M., Ou, M., Ingram, L. O. and Shanmugam, K. T. 2004. "Fermentation of sugar cane bagasse
hemicellulose hydrolysate to l(+)-lactic acid by a thermotolerant acidophilic Bacillus sp.".
Biotechnology Letters 26 (11): 865-868.
Patel, M. A., Ou, M. S., Harbrucker, R., Aldrich, H. C., Buszko, M. L., Ingram, L. O. and Shanmugam, K.
T. 2006. "Isolation and characterization of acid-tolerant, thermophilic bacteria for effective
fermentation of biomass-derived sugars to lactic acid". Applied and Environmental
Microbiology 72 (5): 3228-3235.
Peace, W. R. and Gilani, G. S. 2005. "Chromatographic determination of amino acids in foods". AOAC
International Journal 88 (5): 877–887.
Pelicano, H., Martin, D. S., Xu, R-H and Huang, P., 2006. "Glycolysis inhibition for anticancer
treatment. Oncogene". 25: 4633–4646.
Peng, X., Egashira, T., Hanashiro, K., Masai, E., Nishikawa, S., Katayama, Y., Kimbara, K. and Fukuda,
M. 1998. "Cloning of a Sphingomonas paucimobilis syk-6 gene encoding a novel oxygenase

References
150
that cleaves lignin-related biphenyl and characterization of the enzyme". Applied and
Environmental Microbiology 64 (7): 2520-2527.
Peris-Vicente, J., Simó-Alfonso, E., Gimeno Adelantado, J. V. and Doménech Carbó, M. T. 2005.
"Direct infusion mass spectrometry as a fingerprint of protein-binding media used in works
of art". Rapid Communications in Mass Spectrometry 19 (23): 3463-3467.
Petschacher, B. and Nidetzky, B. 2005. "Engineering Candida tenuis xylose reductase for
improved utilization of NADH: antagonistic effects of multiple side chain replacements
and performance of site-directed mutants under simulated in vivo conditions". Appllied
Environmental Microbiology 71:6390–6393.
PhyloTrac Environmental Sample Analysis. 2011. Website http://www.phylotrac.org/. (Accessed
February 14th, 2011).
Pitkänen, J.-P., Rintala, E., Aristidou, A., Ruohonen, L. and Penttilä, M. 2005. "Xylose chemostat
isolates of Saccharomyces cerevisiae show altered metabolite and enzyme levels compared
with xylose, glucose, and ethanol metabolism of the original strain". Applied Microbiology
and Biotechnology 67 (6): 827-837.
Ramos, C.P., Foster, G., and Collins, M.D. 1997. "Phylogenetic analysis of the genus actinomyces
based on 16S rrna gene sequences: Description of Arcanobacterium phocae sp. nov,
Arcanobacterium bernardiae comb. nov. and Arcanobacterium pyogenes comb. nov."
International Journal of Systematic Bacteriology: 46-53.
Raoult, D., Fournier, P. E. and Drancourt, M. 2004. "What does the future hold for clinical
microbiology?" Nature Reviews Microbiology 2 (2): 151-159.
Reller, L. B., Weinstein, M. P. and Petti, C. A. 2007. "Detection and identification of microorganisms
by gene amplification and sequencing". Clinical Infectious Diseases 44 (8): 1108-1114.
Richard, P., Verho, R., Putkonen, M., Londesborough, J. and Penttila, M. 2003. "Production of
ethanol from l-arabinose by saccharomyces cerevisiae containing a fungal l-arabinose
pathway". FEMS Yeast Res 3: 185 - 189.
Richard, T. 2006. "The effect of lignin on biodegradability". http://www.css.cornell.edu/
compost/calc/lignin.html. (Accessed April 13th, 2009).
Ronald, P., and Admachak, R. 2008. "Tomorrow's table: Organic farming, genetics and the future of
food.” Oxford university press.
Rosenberg, S. L. 1980. "Fermentation of pentose sugars to ethanol and other neutral products by
microorganisms". Enzyme and Microbial Technology 2 (3): 185-193.
Rosselló-Mora, R. and Amann, R. 2001. "The species concept for prokaryotes". Federation of
European Microbiology Societies, Microbiology Reviews 25 (1): 39-67.
Roubroeks, J.P., Andersson, R., Mastromauro, D.I., Christensen, B.E., and Åman, P. 2001. "Molecular
weight, structure and shape of oat (1→3),(1→4)-b-D-glucan fractions obtained by enzymatic
degradation with (1→4)-b-D-glucan 4-glucanohydrolase from Trichoderma
reesei"Carbohydrate Polymers 46 (2001) 275-285.

References
151
Routsias, J. G., Karagounis, P., Parvulesku, G., Legakis, N. J. and Tsakris, A. 2010. "In vitro bactericidal
activity of human [beta]-defensin 2 against nosocomial strains". Peptides 31 (9): 1654-1660.
Rubin, E., M. 2008. "Genomics of cellulosic biofuels.” Nature 454 (14): 841-845.
Sakamoto, K. and Konings, W. N. 2003. "Beer spoilage bacteria and hop resistance". International
Journal of Food Microbiology 89 (2-3): 105-124.
Sakanyan, V., Petrosyan, P., Lecocq, M., Boyaen, A., Legrian, C., Demarez, M., Hallet, N. J. and
Glansdorff, N. 1996. "Genes and enzymes of the acetyl cycle of arginine biosynthesis in
Corynebacterium glutamicum: enzymes evolution in the early steps of the arginine
pathway". Microbiology 142: 99-108.
Sanchez, O., Gasol, J. M., Massana, R., Mas, J. and Pedros-Alio, C. 2007. "Comparison of different
denaturing gradient gel electrophoresis primer sets for the study of marine bacterioplankton
communities". Applied and Environmental Microbiology 73 (18): 5962-5967.
Sanguin, H., Sarniguet, A., Gazengel, K., Moënne-Loccoz, Y. and Grundmann, G. L. 2009.
"Rhizosphere bacterial communities associated with disease suppressiveness stages of take-
all decline in wheat monoculture". New Phytologist 184 (3): 694-707.
Sasaki, M., Adschiri, T. and Arai, K. 2003. "Fractionation of sugarcane bagasse by hydrothermal
treatment". Bioresource Technology 86 (3): 301-304.
Schaal, K., Yassin, A., and Stackebrandt, E. 2006. "The family actinomycetaceae: The genera
actinomyces, actinobaculum, arcanobacterium, varibaculum, and mobiluncus". Prokaryotes
3: 430–537.
Schatz, M. C., Phillippy, A. M., Gajer, P., Desantis, T. Z., Andersen, G. L. and Ravel, J. 2010.
"Integrated microbial survey analysis of prokaryotic communities for the PhyloChip
microarray". Applied and Environmental Microbiology : 303-00310.
Scheffers, D. J. and Pinho, G. M. 2005. "Bacterial cell wall synthesis: new insights from
localization studies". Microbiology and Molecular Biology Reviews 69(4):585–607.
Schuller, M., Sloots, T. P., James, G. S., Halliday, C. L., Carter, I. W. J. and James, G. 2010. "Universal
bacterial identification by PCR and DNA sequencing of 16S rRNA gene". In PCR for clinical
microbiology, 209-214: Springer, Netherlands.
Schuppler, M., Wagner, M., Schon, G. and Gobel, U. B. 1998. "In situ identification of nocardioform
actinomycetes in activated sludge using fluorescent rrna-targeted oligonucleotide probes".
Microbiology 144 (1): 249-259.
Sebat, J. L., Colwell, F. S. and Crawford, R. L. 2003. "Metagenomic profiling: Microarray analysis of an
environmental genomic library". Applied and Environmental Microbiology 69 (8): 4927-4934.
Seiflein, T.A. and Lawrence, J.G. 2006. "Two transsulfurylation pathways in Klebsiella
pneumoniae”. Journal of Bacteriology 188 (1) 65762-5774.
Sessitsch, A., Hackl, E., Wenzl, P., Kilian, A., Kostic, T., Stralis-Pavese, N., Sandjong, B. T. and
Bodrossy, L. 2006. "Diagnostic microbial microarrays in soil ecology". New Phytologist 171
(4): 719-736.

References
152
Shigechi, H., Koh, J., Fujita, Y., Matsumoto, T., Bito, Y., Ueda, M., Satoh, E., Fukuda, H. and Kondo, A.
2004. "Direct production of ethanol from raw corn starch via fermentation by use of a novel
surface-engineered yeast strain codisplaying glucoamylase and alpha-amylase". Applied and
Environmental Microbiology 70 (8): 5037-5040.
Sikander A., Ikrum-ul-Haq, Qadeer, M.A., Iqbal, J. 2002. "Production of citric acid by Aspergillus niger
using cane molasses in a stirred fermentor." Electronic Journal of Biotechnology 5 (3).
Singh, A. and Mishra, P. 1995. "Microbial pentose utilization: Current applications in biotechnology".
Amsterdam: Elsevier.
Siqueira, J.F. Jr. 2003. "Periapical Actinomycosis and infection with Propionibacterium propionicum".
Endodontic Topics 6: 78-95.
Skinner, K. A. and Leathers, T. D. 2004. "Bacterial contaminants of fuel ethanol production". Journal
of Industrial Microbiology and Biotechnology 31 (9): 401-408.
Stackebrandt, E. and Goebel, B. M. 1994. "Taxonomic note: A place for DNA-DNA reassociation and
16S rRNA sequence analysis in the present species definition in bacteriology". International
Journal of Systematic Bacteriology 44 (4): 846-849.
Stanbury P.F., Whitaker A., Hall S.J. 1993 "Principles of fermentation technology". Pergamon.
Stanier, R. Y., Adelberg, E. A. and Ingraham, J. L. eds. 1977. General microbiology. Edited. 4 ed.
London: Macmillan Press.
Steinkraus, K. H. 1995. "Handbook of indigenous fermented foods". New York: Marcel Dekker.
Takai, S.,Narita, K., Ando, K., and S.Tsubaki. 1986.Ecology of Rhodococcus (corynebacterium) equi in
soil on a horse-breeding farm. Veterinary Microbiology, Volume 12, Issue 2; 169–177
Tamang, J.P. and Kailasapathy, K. 2010. "Fermented foods and beverages of the world ". USA: CRC
Press Taylor & Francis Group.
Tang, Y.-W., Von Graevenitz, A., Waddington, M. G., Hopkins, M. K., Smith, D. H., Li, H., Kolbert, C. P.,
Montgomery, S. O. and Persing, D. H. 2000. "Identification of coryneform bacterial isolates
by ribosomal DNA sequence analysis". Journal of Clinical Microbiology 38 (4): 1676-1678.
Tempest, W.D. and Meers, L .J. 1970. "Influence of Environment on the Content and
Composition of Microbial Free Amino Acid Pools". Journal of General Microbiology
64:171-185.
Teusink, B. and Smid, E. J. 2006. "Modelling strategies for the industrial exploitation of lactic acid
bacteria". Nature Reviews Microbiology 4 (1): 46-56.
Teusink, B., Walsh, M. C., Van Dam, K. and Westerhoff, H. V. 1998. "The danger of metabolic
pathways with turbo design". Trends Biochemical Sciences 23: 162 - 169.
Thiele, C., Gänzle, M. G. and Vogel, R. F. 2002. "Sample preparation for amino acid determination by
integrated pulsed amperometric detection in foods". Analytical Biochemistry 310 (2): 171-
178.
The new world encyclopedia. Citric acid cycle,
http://www.newworldencyclopedia.org/entry/Citric_acid_cycle (Access April 4th, 2011).

References
153
Thomson, R., Carter, R., Gilpin, C., Coulter, C. and Hargreaves, M. 2008. "Comparison of methods for
processing drinking water samples for the isolation of Mycobacterium avium and
Mycobacterium intracellulare". Applied and Environmental Microbiology 74 (10): 3094-3098.
Throbäck, I. N., Enwall, K., Jarvis, Å. and Hallin, S. 2004. "Reassessing PCR primers targeting nirs, nirk
and nosz genes for community surveys of denitrifying bacteria with DGGE". FEMS
Microbiology Ecology 49 (3): 401-417.
Todar, K. 2008. "Todar's Online Textbook of Bacteriology, Chapter General Bacteriology, Diversity of
Metabolism in Procaryotes (page 3)". Department of Bacteriology, University of
Wisconsin,1550 Linden Drive, Madison, Wisconsin 53706, USA.
Toivari, M. H., Aristidou, A., Ruohonen, L. and Penttila, M. 2001. "Conversion of xylose to ethanol by
recombinant Saccharomyces cerevisiae: Importance of xylulokinase (xks1) and oxygen
availability". Metabolic Engineering 3: 236 - 249.
Tsukamura, M. 1982. "Numerical analysis of the taxonomy of Nocardiae and Rhodococci". Microbial
Immunology 26 (12): 1101-1119.
Unina (Universita degli Studi di Napoli Federico II). 2009. "What is Chemistry? - Starch".
http://www.whatischemistry.unina.it/en/starch.html (Accessed April 13th, 2009).
U.S. Department of Energy. 2005. "Biofuels joint roadmap". Biomass to Biofuels Workshop,
December 7–9, 2005, Rockville, Maryland.
U.S. National Library of Medicine. The genetics home reference.
http://ghr.nlm.nih.gov/gene/G6PD (Acessed June 2nd, 2011).
Vandamme, P., Pot, B., Gillis, M., De Vos, P., Kersters, K. and Swings, J. 1996. "Polyphasic taxonomy,
a consensus approach to bacterial systematics". Microbiological Reviews 60 (2): 407-438.
Varela, C., Agosin, E., Baez, M., Klapa, M. and Stephanopoulos, G. 2002. "Metabolic flux
redistribution in Corynebacterium glutamicum in response to osmotic stress". Applied
Microbiology and Biotechnology 60 ( 5): 547-555.
Verho, R., Londesborough, J., Penttila, M. and Richard, P. 2003. "Engineering redox cofactor
regeneration for improved pentose fermentation in Saccharomyces cerevisiae". Applied and
Environmental Microbiology 69 (10): 5892-5897.
Von Sivers, M. and Zacchi, G. 1996. "Ethanol from lignocellulosics: A review of the economy".
Bioresource Technology 56: 131 - 140.
Wada, M., Awano, N., Haisa, K., Takagi, H. and Nakamori, S. 2002. "Purification, characterization and
identification of cysteine desulfhydrase of Corynebacterium glutamicum, and its relationship
to cysteine production". Federation of European Microbiological Societies: Microbiology
Letters 217 (1): 103-107.
Wang, L., Hashimoto, Y., Tsao, C.-Y., Valdes, J. J. and Bentley, W. E. 2005. "Cyclic AMP (cAMP) and
cAMP receptor protein influence both synthesis and uptake of extracellular autoinducer 2 in
Escherichia coli". Journal of Bacteriology 187 (6): 2066-2076.

References
154
Watanabe, K. and Kasai, Y. 2008. "Emerging technologies to analyze natural attenuation and
bioremediation". In Microbial biodegradation: Genomics and molecular biology: Norfolk, UK,
Caister Academic Press.
Watanabe, S., Abu, S. A., Pack, S. P., Annaluru, N., Kodaki, T. and Makino, K. 2007."Ethanol
production from xylose by recombinant Saccharomyces cerevisiae expressing protein-
engineered NADH-preferring xylose reductase from Pichia stipitis". Microbiology 153
3044–3054.
Wittmann, C., Krömer, J. O., Kiefer, P., Binz, T. and Heinzle, E. 2004. "Impact of the cold shock
phenomenon on quantification of intracellular metabolites in bacteria". Analytical
Biochemistry 327 (1): 135-139.
Woese C, Kandler O, Wheelis M. 1990. "Towards a natural system of organisms: proposal for the
domains Archaea, Bacteria, and Eucarya.". Proceedings of the National Academy of Science
USA 87 (12): 4576–9.
Womersley, J. 2006. Guideline: Managing impacts from the bulk storage of bagasse. Department of
Environment and Resource Management publication, Queensland, Australia.
Wong, P., Gladney, S. and Keasling, J. D. 1997. "Mathematical model of the lac operon: Inducer
exclusion, catabolite repression, and diauxic growth on glucose and lactose". Biotechnology
Progress 13 (2): 132-143.
Woodward, C., Henderson, W. and Wielgos, T. 2010. "High-speed amino acid analysis (AAA) on sub-
two micron reversed-phase (RP) columns". Pharmaceuticals and Foods, Agilent Technology
Woodward, C. 2007. "Agilent application Note, 5989-6297en".
Xu, H., Dou, W., Xu, H., Zhang, X., Rao, Z., Shi, Z., Xu, Z. 2009."A two-stage oxygen supply strategy
for enhanced l-arginine production by Corynebacterium crenatum based on metabolic
fluxes analysis". Biochemical Engineering Journal 43:41–51.
Yamada, H., Tsukagoshi, N. and Udaka, S. 1981. "Morphological alterations of cell wall concomitant
with protein release in a protein-producing bacterium, Bacillus brevis 47". Journal of
Bacteriology 148 (1): 322-332.
Yanase, H., Sato, D., Yamamoto, K., Matsuda, S., Yamamoto, S. and Okamoto, K. 2007. "Genetic
engineering of Zymobacter palmae for production of ethanol from xylose". Applied and
Environmental Microbiology 73 (8): 2592-2599.
Young, T. K. and Chipley, R J. 1983." Microbial Production of Lysine and Threonine from Whey
Permeate". Applied and Environmental Microbiology 45(2):610-615.
Yu, Z., Garcia-Gonzalez, R., Schanbacher, F. L. and Morrison, M. 2008. "Evaluations of different
hypervariable regions of archaeal 16S rRNA genes in profiling of methanogens by archaea-
specific pcr and denaturing gradient gel electrophoresis". Applied and Environmental
Microbiology 74 (3): 889-893.

References
155
Yusupov, M.M., Yusupova, G.Z., Baucom A., Lieberman, K., Earnest, T.N., Cate, J.H., Noller, H.F.
2001. "Crystal structure of the ribosome at 5.5 A resolution". Science 292 (5518): 883–96.
Zavarzin, G. 2006. "Winogradsky and modern microbiology". Microbiology 75 (5): 501-511.
Zhao, G.-Z., Li, J., Zhu, W.-Y., Klenk, H.-P., Xu, L.-H. and Li, W.-J. 2011. "Nocardia artemisiae sp. Nov.,
an endophytic actinobacterium isolated from surface-sterilized stem of artemisia annual."
International Journal of Systematic and Evolutionary Microbiology 61 (1).
Zhang, Y., Zhu, Y., Zhu, Y., Li, Y. 2010. “The importance of engineeringphysiological
functionality into microbes”. Trends Biotechnol. (In Press).
Zoppa, M. K., Gallo, L., Zacchello, F., Giordano, G. 2006. "Method for the quantification of
underivatized amino acids on dry blood spots from newborn screening by HPLC-ESI-MS/MS
". Journal of Chromatography B 831: 267-273.