Embed Size (px)
Citation preview

DEVELOPMENTS IN FILARIASIS RESEARCH
Lars Volkmann Æ Odile Bain Æ Michael Saeftel
Sabine Specht Æ Kerstin Fischer Æ Frank Brombacher
Klaus I. Matthaei Æ Achim Hoerauf
Murine filariasis: interleukin 4 and interleukin 5 lead to containmentof different worm developmental stages
Received: 22 July 2002 /Accepted: 23 August 2002 / Published online: 10 December 2002� Springer-Verlag 2002
Abstract We compared the impact of IL-4 and IL-5deficiency during the fully permissive infection ofBALB/c mice with the rodent filaria Litomosoidessigmodontis. IL-5, in contrast to IL-4, is crucial for thecontainment of adult worms during short- and long-term infections. IL-5 KO mice allowed development ofmore larvae into adult worms and showed up to 200times more adult worms persisting during chronic in-fection (day 60 until 200 post-infection). This increasedpersistence was accompanied by a reduction in inflam-matory nodules around adult filariae. In contrast, adultworm survival and nodule formation did not differ be-tween BALB/c wild-type mice and BALB/c IL-4 KO orBALB/c IL-4 receptor (IL-4R) a-chain KO mice. Inboth IL-4 and IL-5 KO mice microfilaraemia wasgreatly enhanced (160-fold) and prolonged compared towild-type mice. This extent of susceptibility to microfil-ariae required the presence of adult worms in a full in-fection cycle since upon intraperitoneal injection ofmicrofilariae alone they were removed from BALB/c,BALB/c IL-4 KO and BALB/c IL-4R a-chain KO mice
with equivalent kinetics, and since microfilarial survivalwas only slightly increased in IL-5 KO mice (factor of 5vs. factor of 160 in full infection). In conclusion, IL-4and IL-5 dependent effector pathways operate againstdifferent stages of filarial worms, and IL-5 has a greaterimpact on parasite containment than IL-4.
Keywords Interleukin-4 Æ Interleukin-5 Æ Murinefilariasis Æ Neutrophils Æ Eosinophils
Introduction
Filariasis is a major social and economic burden in thetropics and sub-tropics of Asia, Africa, western Pacificand part of the Americas, striking over 180 millionpersons in 73 countries. Mammalian hosts are infectedduring bloodmeals of arthropods which transmit infec-tive third stage larvae (L3). L3 mature, molting twice, toadult worms which release microfilariae (MF) into eitherskin or blood.
To identify the immune reactions which are essentialfor parasite control the infection model of BALB/cmice with the rodent filarial species Litomosoidessigmodontis was established [33]. This model is con-sidered advantageous since it permits the monitoring ofL3 development into adult worms and the adult wormpersistence during patency. The full permissivity for theparasite in immunocompetent laboratory mice allowsthe examination of immunity against all developmentalstages of the parasite during a course of infection. Inprevious studies using this model we showed that bothTh1 and Th2 immune responses are involved in resis-tance to filarial infections. Interferon (IFN) c deficiencyresulted in a higher worm survival due to reducedformation of nodules around adult filariae [34]. Asimilarly impaired nodule formation and a highermicrofilaraemia were observed after neutralisation ofinterleukin (IL) 5 [2]. IL-5 is also essential for vaccine-induced protection: in vaccinated mice eosinophils arequickly recruited to the subcutaneous tissue after
Med Microbiol Immunol (2003) 192: 23–31DOI 10.1007/s00430-002-0155-9
L. Volkmann Æ M. Saeftel Æ S. Specht Æ K. FischerA. Hoerauf (&)Bernhard Nocht Institute for Tropical Medicine, Hamburg,Germany, Bernhard Nocht Strasse 74,20359 Hamburg, GermanyE-mail: [email protected].: +49-40-42818301Fax: +49-40-42818400
O. BainInstitut de Systematique CNRS FR 1541,Biologie Parasitaire, Museum National d’Histoire Naturelle,Paris, France
F. BrombacherDepartment of Immunology, Health Faculty,Groote Schuur Hospital, University of Cape Town,Cape Town, South Africa
K.I. MatthaeiDivision of Biochemistry and Molecular Biology,John Curtin School of Medical Research,Australian National University Canberra, Australia

challenge with L3, and larvae are destroyed bydegranulation and release of toxic products. The re-cruitment of eosinophils is dependent on IL-5 [23,25]while the eosinophil degranulation is abolished in theabsence of B cells, suggesting a role for antibodies ineosinophil degranulation [27]. In contrast, in primaryinfection during patency (production of MF), effectormechanisms for both IL-5 and IFN-c apparently in-volve the production of tumor necrosis factor (TNF) aand KC (the murine homologue of IL-8) which areassociated with recruitment of neutrophils [2,34].Neutrophils have been shown to be essential for nor-mal worm killing [2]. In contrast, the defect in IL-4production did not lead to a higher survival of adultworms, but it led to highly enhanced MF loads [35].
IL-4R a chain KO mice allow the monitoring ofIL-13 and IL-4 driven effector functions [7,37] Thesemice have significant levels of IL-13 and IL-4, but theeffector functions of both cytokines are abrogated[11,30] In an infection system with Schistosoma mansoniprotective immune responses were not dependent onIL-4 when IL-4 deficient C57BL/6 mice were used[14,18] but on IL-13, as shown in BALB/c IL-4R a chainKO [29].
In this study we compared the course of L. sigm-odontis infection in BALB/c mice, IL-5 KO, IL-4 KO orBALB/c IL-4 KO mice, and BALB/c IL-4R a chain KO.We focused on the comparison of parasite developmentin the early stage of infection and the outcome of theinfection through the whole period of patency. While IL-4 and IL-4R deficiency lead to a massive impact on thesurvival of only one stage, the MF, IL-5 has an impacton the survival of all developmental stages of the para-site in the murine host.
Material and methods
Animal maintenance and infection of mice with L. sigmodontis
BALB/c IL-4<tm2Nt> (backcrossed from C57BL/6 to BALB/cbackground – hereafter IL-4 KO) were obtained from the JacksonLaboratory (Bar Harbor, Maine, USA). BALB/c mice withtargetted disruptions of the IL-4 [31] (hereafter BALB/c IL-4 KOmice) and IL-4R a gene [28] (hereafter BALB/c IL-4Ra KO)derived from BALB/c embryonal stem cells were a kind of giftfrom F. Brombacher (Groote Schuur Hospital, University ofCape Town, Cape Town, South Africa). IL-5 KO mice [19] weresupplied by K.I.M.. The IL-5 KO was generated in C67BL/6embryonal stem cells and then crossed to BALB/c for ten gene-rations.
The KO strains were housed under specific pathogen-freeconditions in micro-isolator cages. KO offspring and BALB/cwild-type mice (originally from Charles River, Sulzfeld, Germany)and cotton rats were bred at the animal facilities of the BernhardNocht Institute. Natural infections of mice with L. sigmodontiswere performed as described previously [1,3]. L. sigmodontis livesnaturally in the cotton rat (Sigmodon hispidus) which can survivevery high blood MF levels (up to 10000 MF/ll blood). During theblood meal on an infected rat the arthropod intermediate host(Ornithonyssus bacoti mite) ingests the MF which molt twice anddevelop to the infective stage 3 larvae (L3) within 10 days. In afollowing blood meal the mites transmit the L3 onto cotton rats.
Similarly, in the case of murine infection the L3 containing mitesare allowed to take blood from mice and thereby transmit the L3.In the rodent host the L3 migrate to the thoracic cavity and reachsexual maturity within 25–33 days. Viviparous female worms startproducing MF which are detectable in the blood after day 50post-infection (p.i.). At least four mice were used for each treat-ment group.
Parasite and inflammatory nodule recovery
The number of adult worms was counted at days 28, 60, 80, 111,130 and 200 days p.i.. To this, the thoracic cavity, the representa-tive site for assessment of worm numbers [3,24] was flushed with10 ml phosphate-buffered solution (PBS)/1% fetal calf serum(FCS), and worms were allowed to sediment. The sediment wasalso used to determine the number of inflammatory nodules.Microfilaraemia was determined in EDTA-treated peripheral bloodafter staining with Hinkelmann’s solution (0.5% w/v eosin Y, 0.5%w/v phenol and 0.185 v/v formaldehyde in distilled water), as de-scribed [3]. For the examination of uterine MF loads adult femaleworms were cleared in lactophenol and examined under themicroscope (magnifications of ·20–100).
Purification of peripheral blood MF
MF were purified from the peripheral blood of cotton rats on aPercoll gradient as described [8] Iso-osmotic Percoll (IOP) wasprepared by mixing nine parts of Percoll (density 1.130 g/ml) withone part of 2.5 M sucrose. Various dilutions of the IOP in 0.25 Msucrose were made to obtain 25%, 30% and 35% solutions. Thegradient dilutions with 2 ml of each dilution were layered in 15-mltubes, and then 2 ml of two-fold diluted peripheral blood waspipetted on the top of the P layers. The tubes were then centrifugedat 400 g for 30 min at room temperature. Recovered MF (betweenthe 25% and 30% layers) were washed with RPMI 1460 1% FCS,and 150,000 viable MF were injected intraperitoneally into naivemice.
MF recovery from the peritoneal cavity
The number of MF in the peritoneal cavity was counted 7, 14 and21 days after the intraperitoneal injection. To this the peritonealcavity was flushed with 10 ml PBS/1% FCS. MF burdens wereexamined after staining with Hinkelmann’s solution.
Determination of proportions of inflammatory cellsin thoracic cavity fluid
Proportions of inflammatory, pleural exudate cells (PLECs) weredetermined using cytospin preparations: 200 ll thoracic cavity cells(2·105/ml in PBS/1% FCS) were centrifuged against a glass slidewith absorptive filter paper using a Shandon cytocentrifuge.Cytospins were then stained using the Wright-Giemsa stain (Sigma,Munich, Germany), and neutrophils, eosinophils, macrophagesand lymphocytes were differentially enumerated.
Flow cytometric analysis
Proportions of B1 and B2 cells in the thoracic cavity were deter-mined using Flow cytometric analysis. We incubated 100 ll tho-racic cavity cells (2 ·106/ml in RPMI 1460 1% FCS) for 30 min onice with a combination of 1:100 dilution of anti-B220-PE (RA3–6B2) and anti-Mac-1–fluorescein isothiocyanate (M1/170.15). Thecells were washed with PBS 1% bovine serum albumin and fixed in1% (v/v) formaldehyde. The samples were measured by a FACScan(Becton Dickinson, Heidelberg, Germany) and analysed by usingCell Quest software (Becton Dickinson). Viable lymphocytes were
24

gated with the forward and side scatter, and 5000 gated events werecollected.
Cell culture
The culture of thoracic cavity MPs was carried out after adhesionon 96-well culture plates (Greiner, Frickenkausen, Germany) inRPMI/5% FCS at 37�C and 5% CO2 for 2 h. We first performed aFACS analysis to determine the relative proportion of MP in thethoracic cavity cells. According to these data, the cell input wascalculated such that 40,000 macrophages/well were allowed to ad-here. The non-adherent cells were removed by washing three timeswith PBS. Purity was greater than 95% (not shown). MP werecultured for 24 h in the presence of only RPMI 1460 5% FCS,100 lg/ml adult worm antigen or with 20 ng/ml IFN-c (Pharmin-gen, Heidelberg, Germany) and 20 ng/ml lipopolysaccharide(Escherichia coli O55, Sigma, Munich, Germany). Supernatantswere then removed for cytokine determination.
Cytokine assays
Concentrations of cytokines IL-5, IL-10 and TNF-a in pleura orperitoneal exudates (thoracic and peritoneal wash) were determinedby specific two-site enzyme-linked immunosorbent assays (ELISAs)using standard protocols. The antibody pairs for capture and de-tection (biotinylated) were purchased from Pharmingen in thecombination recommended. Recombinant cytokines (Pharmingenand R&D Systems, Wiesbaden, Germany) were used as standards.Mouse KC (homologue of human IL-8) was determined using af-finity purified goat IgG as capture (R&D, cat. no. MAB453) and asbiotinylated detector antibody (cat. no. BAF453). Recombinantchemokine (cat. no. 453-KC-010) was used as standard. All ELI-SAs were developed after incubation with streptavidin-peroxidasecomplex (1:10000; Boehringer-Mannheim, Mannheim, Germany),using 3,5,3¢,5¢ tetramethylbenzidine as substrate (Roth, Karlsruhe,Germany; dissolved 6 mg/ml in DMSO). Sensitivity was 10 pg/ml.
Immunoglobulin assays
Phosphorylcholine (PC) specific serum antibodies were quantifiedby coating Nunc Polysorb microtitre plates (Nunc, Roskilde,Denmark) with 10 mg/ml phosphorylcholine–bovine serum albu-min. After incubation with sera and washing (using standard pro-
cedures), plates were incubated with biotinylated antimurine IgMmonoclonal antibody (clone II/41; PharMingen) or with anti-mouse IgG antibodies (The Binding Site, Heidelberg, Germany).All ELISAs were developed as described for cytokine ELISAs.
Statistical analyses
The nonparametric Mann-Whitney U test was used to comparefilarial recovery rates, number of inflammatory cells, cytokines andimmunoglobulins levels. P values less than 0.05 were considered toindicate a statistically significant difference.
Results
IL-5 deficiency but not IL-4 and IL-4R a deficiencyincreases susceptibility to L. sigmodontis adult worms
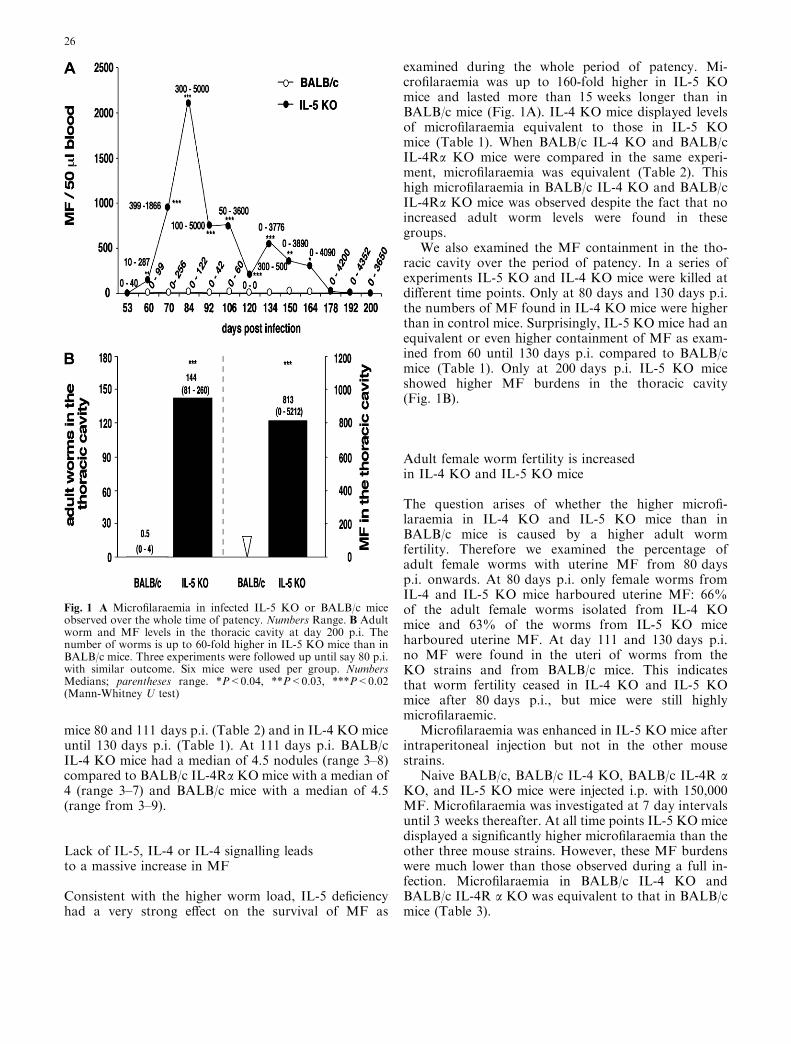
IL-5 deficiency had a significant effect on the adult wormload 28 days p.i. (Table 1). This indicates that the lackof IL-5 influenced the development from L3 to adultworms, i.e. worm establishment. In addition, IL-5 defi-ciency drastically enhanced adult worm survival at thelate stages of infection as observed from day 60 until200 days p.i. (Table 1, Fig. 1B). In contrast, in BALB/cand IL-4 KO mice adult worm loads decreased from60 days p.i. to 130 days p.i. to almost absent levels(Table 1). IL-5 KO mice showed lower worm encapsu-lation than IL-4 KO and BALB/c mice. IL-5 KO micehad a median of 1.5 nodules (range 0–4) 80 days p.i.compared to IL-4 KO mice, with a median of 4 (range 3–7) and BALB/c mice with a median of 5 (range 4–7;P <0.05, Mann-Whitney U test). At 200 days p.i., whilethe medians of adult worm loads in BALB/c mice werezero, IL-5 KO mice still had high parasite burdens(Fig. 1B). Neither IL-4 nor IL-4 signalling was essentialfor adult worm killing in the late stage of infection asexamined in BALB/c IL-4 KO and BALB/c IL-4Ra KO
Day p.i. Adult worms MF in 50 ll peripheral blood MF in the thoracic cavity
BALB/c IL-4 KOa IL-5 KO BALB/c IL-4 KO IL-5 KO BALB/c IL-4 KO IL-5 KO
28 39 (9–54) 52 (19–56) 55* (43–130) n.d. n.d. n.d. n.d. n.d. n.d.60 22 (8–39) 68 (28–87) 175.5**
(105–253)6 (0–120) 128*
(0–1,238)100*(0–3,021)
6,8314*(3,311–7,111)
77,7504*(3,411–273,315)
150(0–1014)
80 14 (9–27) 41 (26–50) 121**(90–235)
2 (0–44) 548***(178–6,426)
291***(0–1,272)
2,521(1,102–4,213)
285,0525*(140,321–870,055)
101(0–789)
111b 0 7 99 0 (0–3) 1092*(46–3,392)
2952*(140–5,746)
1,185 895 1211
130 1.5 (1–2) 2.5 (0–5) 127**(105–231)
2 (0–4) 1460.5*(112–2,809)
2279*(445–4,654)
50 (0–100) 2,845*(1,102–4,213)
1,423*(105–2,421)
*P<0.04 vs. BALB/c mice, **P<0.04 vs. BALB/c and IL-4 KO mice, ***P<0.01 vs. BALB/c mice (MF were counted in seven BALB/cmice and eight IL-4 KO mice), 4*P<0.04 vs. IL-5 KO mice, 5*P<0.04 vs. BALB/c mice and IL-5 KO miceaIL-4 KO mice were backcrossed from C57BL/6 to BALB/c backgroundbAt 111 days p.i. only one mouse per group was analysed for adult worm and thoracic MF load detection. In all other cases if not noteddifferently four mice were used per group
Table 1 Parasite loads of adult worms and MF in the peripheralblood and in the thoracic cavity in BALB/c, IL-4 KO and IL-5 KOmice. Difference in worm load between different time points reflectsthe different infection loads in these experiments; three experiments
were followed up until 80 days p.i. with similar outcome. Valuesgiven are medians (range); statistical analysis used the Mann-Whitney U test
25
mice 80 and 111 days p.i. (Table 2) and in IL-4 KO miceuntil 130 days p.i. (Table 1). At 111 days p.i. BALB/cIL-4 KO mice had a median of 4.5 nodules (range 3–8)compared to BALB/c IL-4Ra KO mice with a median of4 (range 3–7) and BALB/c mice with a median of 4.5(range from 3–9).
Lack of IL-5, IL-4 or IL-4 signalling leadsto a massive increase in MF
Consistent with the higher worm load, IL-5 deficiencyhad a very strong effect on the survival of MF as
examined during the whole period of patency. Mi-crofilaraemia was up to 160-fold higher in IL-5 KOmice and lasted more than 15 weeks longer than inBALB/c mice (Fig. 1A). IL-4 KO mice displayed levelsof microfilaraemia equivalent to those in IL-5 KOmice (Table 1). When BALB/c IL-4 KO and BALB/cIL-4Ra KO mice were compared in the same experi-ment, microfilaraemia was equivalent (Table 2). Thishigh microfilaraemia in BALB/c IL-4 KO and BALB/cIL-4Ra KO mice was observed despite the fact that noincreased adult worm levels were found in thesegroups.
We also examined the MF containment in the tho-racic cavity over the period of patency. In a series ofexperiments IL-5 KO and IL-4 KO mice were killed atdifferent time points. Only at 80 days and 130 days p.i.the numbers of MF found in IL-4 KO mice were higherthan in control mice. Surprisingly, IL-5 KO mice had anequivalent or even higher containment of MF as exam-ined from 60 until 130 days p.i. compared to BALB/cmice (Table 1). Only at 200 days p.i. IL-5 KO miceshowed higher MF burdens in the thoracic cavity(Fig. 1B).
Adult female worm fertility is increasedin IL-4 KO and IL-5 KO mice
The question arises of whether the higher microfi-laraemia in IL-4 KO and IL-5 KO mice than inBALB/c mice is caused by a higher adult wormfertility. Therefore we examined the percentage ofadult female worms with uterine MF from 80 daysp.i. onwards. At 80 days p.i. only female worms fromIL-4 and IL-5 KO mice harboured uterine MF: 66%of the adult female worms isolated from IL-4 KOmice and 63% of the worms from IL-5 KO miceharboured uterine MF. At day 111 and 130 days p.i.no MF were found in the uteri of worms from theKO strains and from BALB/c mice. This indicatesthat worm fertility ceased in IL-4 KO and IL-5 KOmice after 80 days p.i., but mice were still highlymicrofilaraemic.
Microfilaraemia was enhanced in IL-5 KO mice afterintraperitoneal injection but not in the other mousestrains.
Naive BALB/c, BALB/c IL-4 KO, BALB/c IL-4R aKO, and IL-5 KO mice were injected i.p. with 150,000MF. Microfilaraemia was investigated at 7 day intervalsuntil 3 weeks thereafter. At all time points IL-5 KO micedisplayed a significantly higher microfilaraemia than theother three mouse strains. However, these MF burdenswere much lower than those observed during a full in-fection. Microfilaraemia in BALB/c IL-4 KO andBALB/c IL-4R a KO was equivalent to that in BALB/cmice (Table 3).
Fig. 1 A Microfilaraemia in infected IL-5 KO or BALB/c miceobserved over the whole time of patency. Numbers Range. B Adultworm and MF levels in the thoracic cavity at day 200 p.i. Thenumber of worms is up to 60-fold higher in IL-5 KO mice than inBALB/c mice. Three experiments were followed up until say 80 p.i.with similar outcome. Six mice were used per group. NumbersMedians; parentheses range. *P<0.04, **P<0.03, ***P<0.02(Mann-Whitney U test)
26
IL-5 deficient mice have a significantly reducedaccumulation of B1 cells in the thoracic cavityand significantly reduced PC-specific Ig M-antibodylevels in the serum
B1 cells and IgM antibodies were demonstrated to beimportant for the control of L3 to adult wormdevelopment [1]. IL-5 KO mice in this study showed alower accumulation of B1 cells and B2 cells into thethoracic cavity 28 days p.i. than BALB/c and IL-4 KOmice (Table 4). To assess whether the lower number B1cells in IL-5 KO is associated with a lower immuneresponse against L3 and L4 larvae we determined thehumoral response in this stage of infection and innoninfected mice. The examination of PC-specific IgM-antibodies 0 and 28 days p.i. revealed significantly lowerlevel in IL-5 KO mice than in IL-4 KO mice or BALB/cmice (Table 4).
Eosinophil, neutrophil and neutrophil chemotacticcytokines are diminished in IL-5 deficient mice
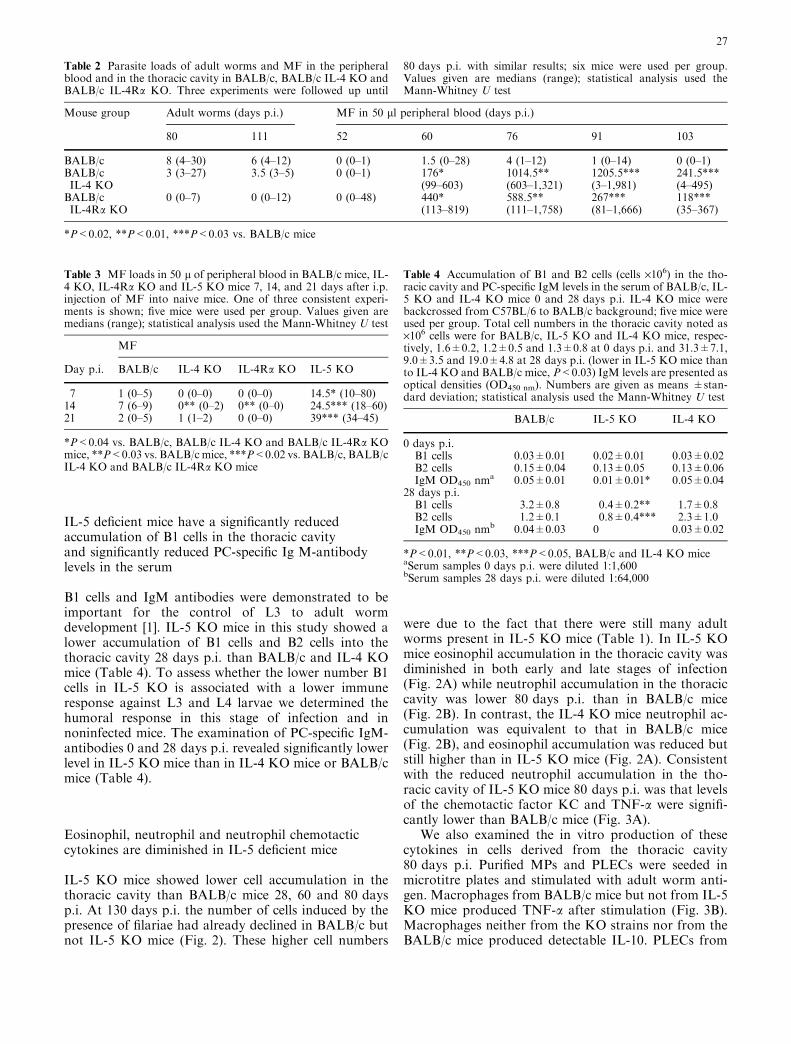
IL-5 KO mice showed lower cell accumulation in thethoracic cavity than BALB/c mice 28, 60 and 80 daysp.i. At 130 days p.i. the number of cells induced by thepresence of filariae had already declined in BALB/c butnot IL-5 KO mice (Fig. 2). These higher cell numbers
were due to the fact that there were still many adultworms present in IL-5 KO mice (Table 1). In IL-5 KOmice eosinophil accumulation in the thoracic cavity wasdiminished in both early and late stages of infection(Fig. 2A) while neutrophil accumulation in the thoraciccavity was lower 80 days p.i. than in BALB/c mice(Fig. 2B). In contrast, the IL-4 KO mice neutrophil ac-cumulation was equivalent to that in BALB/c mice(Fig. 2B), and eosinophil accumulation was reduced butstill higher than in IL-5 KO mice (Fig. 2A). Consistentwith the reduced neutrophil accumulation in the tho-racic cavity of IL-5 KO mice 80 days p.i. was that levelsof the chemotactic factor KC and TNF-a were signifi-cantly lower than BALB/c mice (Fig. 3A).
We also examined the in vitro production of thesecytokines in cells derived from the thoracic cavity80 days p.i. Purified MPs and PLECs were seeded inmicrotitre plates and stimulated with adult worm anti-gen. Macrophages from BALB/c mice but not from IL-5KO mice produced TNF-a after stimulation (Fig. 3B).Macrophages neither from the KO strains nor from theBALB/c mice produced detectable IL-10. PLECs from
Mouse group Adult worms (days p.i.) MF in 50 ll peripheral blood (days p.i.)
80 111 52 60 76 91 103
BALB/c 8 (4–30) 6 (4–12) 0 (0–1) 1.5 (0–28) 4 (1–12) 1 (0–14) 0 (0–1)BALB/cIL-4 KO
3 (3–27) 3.5 (3–5) 0 (0–1) 176*(99–603)
1014.5**(603–1,321)
1205.5***(3–1,981)
241.5***(4–495)
BALB/cIL-4Ra KO
0 (0–7) 0 (0–12) 0 (0–48) 440*(113–819)
588.5**(111–1,758)
267***(81–1,666)
118***(35–367)
*P<0.02, **P<0.01, ***P<0.03 vs. BALB/c mice
Table 2 Parasite loads of adult worms and MF in the peripheralblood and in the thoracic cavity in BALB/c, BALB/c IL-4 KO andBALB/c IL-4Ra KO. Three experiments were followed up until
80 days p.i. with similar results; six mice were used per group.Values given are medians (range); statistical analysis used theMann-Whitney U test
Table 3 MF loads in 50 l of peripheral blood in BALB/c mice, IL-4 KO, IL-4Ra KO and IL-5 KO mice 7, 14, and 21 days after i.p.injection of MF into naive mice. One of three consistent experi-ments is shown; five mice were used per group. Values given aremedians (range); statistical analysis used the Mann-Whitney U test
MF
Day p.i. BALB/c IL-4 KO IL-4Ra KO IL-5 KO
7 1 (0–5) 0 (0–0) 0 (0–0) 14.5* (10–80)14 7 (6–9) 0** (0–2) 0** (0–0) 24.5*** (18–60)21 2 (0–5) 1 (1–2) 0 (0–0) 39*** (34–45)
*P<0.04 vs. BALB/c, BALB/c IL-4 KO and BALB/c IL-4Ra KOmice, **P<0.03 vs. BALB/c mice, ***P<0.02 vs. BALB/c, BALB/cIL-4 KO and BALB/c IL-4Ra KO mice
Table 4 Accumulation of B1 and B2 cells (cells ·106) in the tho-racic cavity and PC-specific IgM levels in the serum of BALB/c, IL-5 KO and IL-4 KO mice 0 and 28 days p.i. IL-4 KO mice werebackcrossed from C57BL/6 to BALB/c background; five mice wereused per group. Total cell numbers in the thoracic cavity noted as·106 cells were for BALB/c, IL-5 KO and IL-4 KO mice, respec-tively, 1.6±0.2, 1.2±0.5 and 1.3±0.8 at 0 days p.i. and 31.3±7.1,9.0±3.5 and 19.0±4.8 at 28 days p.i. (lower in IL-5 KO mice thanto IL-4 KO and BALB/c mice, P<0.03) IgM levels are presented asoptical densities (OD450 nm). Numbers are given as means ±stan-dard deviation; statistical analysis used the Mann-Whitney U test
BALB/c IL-5 KO IL-4 KO
0 days p.i.B1 cells 0.03±0.01 0.02±0.01 0.03±0.02B2 cells 0.15±0.04 0.13±0.05 0.13±0.06IgM OD450 nm
a 0.05±0.01 0.01±0.01* 0.05±0.0428 days p.i.B1 cells 3.2±0.8 0.4±0.2** 1.7±0.8B2 cells 1.2±0.1 0.8±0.4*** 2.3±1.0IgM OD450 nm
b 0.04±0.03 0 0.03±0.02
*P<0.01, **P<0.03, ***P<0.05, BALB/c and IL-4 KO miceaSerum samples 0 days p.i. were diluted 1:1,600bSerum samples 28 days p.i. were diluted 1:64,000
27
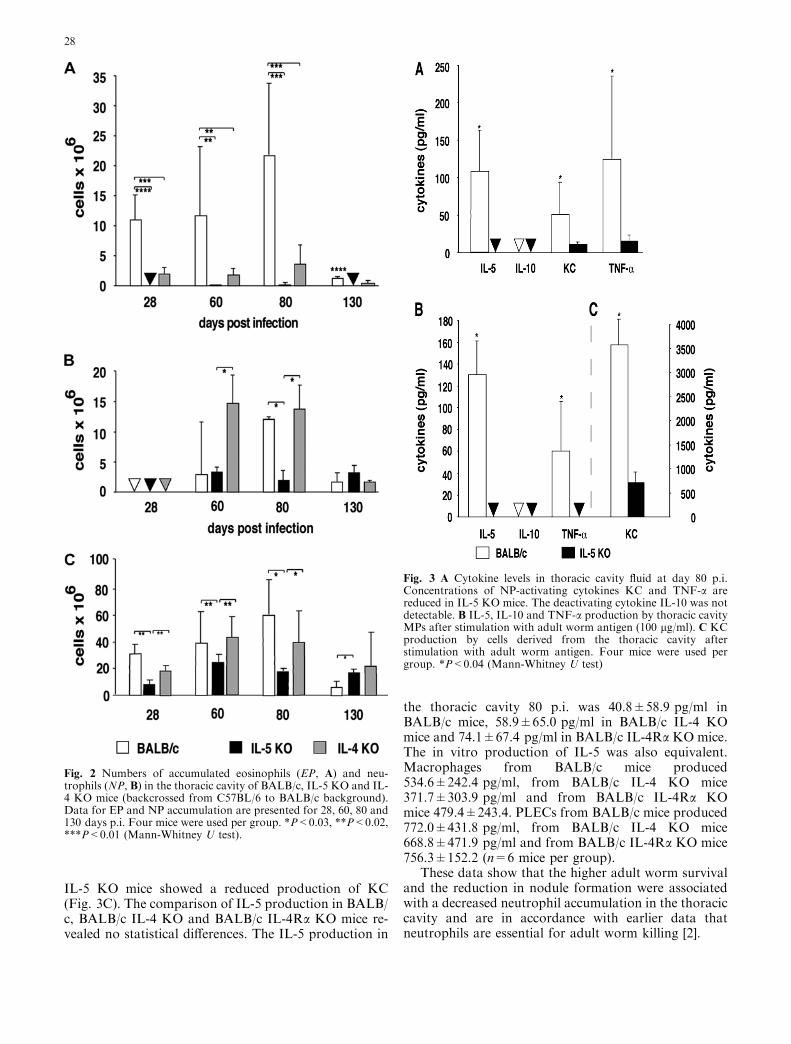
IL-5 KO mice showed a reduced production of KC(Fig. 3C). The comparison of IL-5 production in BALB/c, BALB/c IL-4 KO and BALB/c IL-4Ra KO mice re-vealed no statistical differences. The IL-5 production in
the thoracic cavity 80 p.i. was 40.8±58.9 pg/ml inBALB/c mice, 58.9±65.0 pg/ml in BALB/c IL-4 KOmice and 74.1±67.4 pg/ml in BALB/c IL-4Ra KO mice.The in vitro production of IL-5 was also equivalent.Macrophages from BALB/c mice produced534.6±242.4 pg/ml, from BALB/c IL-4 KO mice371.7±303.9 pg/ml and from BALB/c IL-4Ra KOmice 479.4±243.4. PLECs from BALB/c mice produced772.0±431.8 pg/ml, from BALB/c IL-4 KO mice668.8±471.9 pg/ml and from BALB/c IL-4Ra KO mice756.3±152.2 (n=6 mice per group).
These data show that the higher adult worm survivaland the reduction in nodule formation were associatedwith a decreased neutrophil accumulation in the thoraciccavity and are in accordance with earlier data thatneutrophils are essential for adult worm killing [2].
Fig. 2 Numbers of accumulated eosinophils (EP, A) and neu-trophils (NP, B) in the thoracic cavity of BALB/c, IL-5 KO and IL-4 KO mice (backcrossed from C57BL/6 to BALB/c background).Data for EP and NP accumulation are presented for 28, 60, 80 and130 days p.i. Four mice were used per group. *P<0.03, **P<0.02,***P<0.01 (Mann-Whitney U test).
Fig. 3 A Cytokine levels in thoracic cavity fluid at day 80 p.i.Concentrations of NP-activating cytokines KC and TNF-a arereduced in IL-5 KO mice. The deactivating cytokine IL-10 was notdetectable. B IL-5, IL-10 and TNF-a production by thoracic cavityMPs after stimulation with adult worm antigen (100 lg/ml). C KCproduction by cells derived from the thoracic cavity afterstimulation with adult worm antigen. Four mice were used pergroup. *P<0.04 (Mann-Whitney U test)
28

Discussion
It is accepted that Th2 cytokines are key mediators of anantifilarial immune response and act to limit parasiteloads in the hosts [12,21]. This study contains novel datashowing that the Th2 cytokines IL-4, IL-5 and IL-13control different parasite developmental stages. BALB/cIL-4 KO, BALB/c IL-4Ra KO and BALB/c miceshowed a similar containment of the adult worm loads(Table 2). In contrast, there was significantly lesscontainment of the worm loads in infected IL-5 KOduring worm development, and adult worm persistence(Table 1, Fig. 1B). IL-4 KO or BALB/c IL-4 KO,BALB/c IL-4Ra KO and IL-5 KO mice developed ahighly increased and extended microfilaraemia com-pared to BALB/c mice (Table 2, Fig. 1A). In a previousstudy we showed that in an IL-4 deficient system femaleworm fertility was increased [35]. Here we provide novelinformation that this influence is mediated through IL-4signalling, because BALB/c IL-4Ra KO mice did notshow lower levels of MF in the blood, suggesting thatIL-4 does not act directly on the worm to limit fertility.In addition, the finding that BALB/c IL-4Ra KO,BALB/c IL-4 KO or IL-4 KO mice showed equivalentadult worm loads suggests that IL-13 has no additionaleffects over IL-4 in parasite containment in this systemsince IL-13 acts via receptor signalling of the IL-4Ra-chain [28]. Also the experiments revealed that IL-13had no effect on the control of MF and adult worms inthis system (Table 2). Thus the role of IL-13 in murinefilarial infections is different from that observed inS. mansoni infected mice, in which IL-13 seems to beinvolved in granuloma formation [16] and also wormcontainment [18,29] independently of IL-4.
This study revealed contrasting roles of IL-5 vs. IL-4and IL-4Ra KO regarding the generation of an effectiveimmune response against adult filariae. In BALB/c mice,as well as in IL-4 KO, BALB/c IL-4 KO and BALB/cIL-4Ra KO mice the immune response to adult wormsduring patency (starting 60 days p.i.) was characterisedby the cellular encapsulation of the parasites. This pro-cess has been described to lead to the formation of in-flammatory nodules and to the death of adult worms[2,34]. IL-5 deficient mice, in contrast, showed almost noreduction of the adult worm loads over the whole ob-servation period (Table 1). In a former study we neu-tralised IL-5 by the injection of anti-IL-5 antibodies [2].Compared to the former study, IL-5 KO mice had sig-nificantly higher adult worm loads and microfilaraemiathan the IL-5 neutralised mice [2]. Thus one can con-clude that the neutralisation of the cytokine is less ef-fective than the genetic deficiency, although similarmechanisms of defense may be inhibited.
During the present study we detected no accumula-tion of eosinophils in the thoracic cavity of IL-5 KOmice (Fig. 2A). In helminthic infections IL-5 and eoino-phils are known to be essential for the generation of aneffective immune response [9]. In other nonpermissive
models of filarial infections eosinophils are importanteffector cells for the containment of MF [10] L3, pre-adult stages [5, 20,36] and of adult worms [32]. In theL. sigmodontis infection model it has been hypothesisedthat eosinophils have different roles in vaccination thanin primary infections.
In vaccinated mice there is an accumulation of eo-sinophils in the subcutaneous tissue and thus, a fewhours after the L3 have invaded the subcutaneous tissuethey are attacked by eosinophils [22, 23,25] and arekilled through antibody-dependent cellular cytotoxicity[27]. In contrast, in primary infection it is believed thateosinophils accumulate too late in the subcutaneoustissue, i.e. after L3 have already migrated from the skinto the thoracic cavity [4]. Consistent with these obser-vations there was no reduced worm killing in primaryinfection in IL-5 neutralised mice compared to BALB/cmice in the early phase of infection (28 days p.i.) [2,25].However, the present data reveal that the genetic defectof IL-5, different to that with IL-5 neutralisation [2],does alter the development from L3 into adult wormscompared to BALB/c mice, since in IL-5 KO mice, butnot in IL-5 neutralised mice, there was an greater wormload than in BALB/c 28 days p.i. This may be due to arole of eosinophils which may be more thoroughly re-duced in numbers in IL-5 KO mice as than in anti-IL-5treated mice [2]. Another explanation could be the re-duced levels of PC-specific IgM-antibodies at the crucialinitial days of infection [3,24] in IL-5 KO mice (Table 4)but not in anti-IL-5 treated mice since the anti-IL-5treatment was started only a few days before infectionand could not have affected levels of preformed naturalantibodies. The lack of anti-PC-specific IgM antibodieshas been associated with higher adult worm develop-ment as shown in L. sigmodontis infected BALB.Xidmice [1].
In contrast to eosinophils, neutrophils are not in-volved in the immune response in the early phase ofinfection [26] but in the late stages of infection [2] whenfemale worms produce MF. In the present study weshowed that neutrophil accumulation and nodule for-mation in IL-5 KO mice were significantly lower than inBALB/c mice as observed 80 days p.i. (Fig. 2B) Earlierit was shown by neutrophil depletion in vivo using an-tibodies to granulocyte colony-stimulating factor(G-CSF) that neutrophils are essential for nodule for-mation and adult worm attack [2]. It was then suggestedthat the action of IL-5 on adult worms was indirect,involving neutrophil chemotactic cytokines TNF-a, KCand G-CSF which were produced at much lower levelsin anti-IL-5 treated mice [2]. The present data using IL-5KO mice clearly confirm this view (Fig. 3). In additionto showing that the genetic defect is more effective thanIL-5 neutralisation, the new data on IL-5 KO mice alsorule out that some of the earlier observations may havebeen artefacts induced by prolonged monoclonal anti-body (control or anti-IL-5) application. BALB/c IL-4KO and BALB/c IL-4Ra KO mice did not showreduced IL-5 levels in the thoracic cavity and is in
29

accordance with earlier experiments in which IL-5 wasshown to be regulated independently of IL-4 in filarialinfections [17,35].
Interestingly, also in the human system macrophagesproduced neutrophil chemotactic cytokines in responseto MF producing Onchocerca volvulus worms [6]. Thisproduction is dependent on the presence of Wolbachiaendobacteria which are true endosymbionts essential forworm fertility [13]. Also in the Onchocerca nodule con-taining the adult worm, neutrophil presence is depen-dent on the Wolbachia [6].
The highly increased and extended microfilaraemia inIL-5 KO and IL-4 KO mice (Fig. 1A, Tables 1, 2) wasdue to a significantly extended fertility period of femalefilariae compared to BALB/c mice. Fertility in the defi-cient mice ceased between 80 and 111 days p.i. Never-theless, IL-5 KO, IL-4 KO or BALB/c IL-4 KO andBALB/c IL-4Ra KO mice remained microfilaraemic formany weeks after day 80 p.i. The latter is probably dueto immunosuppressive effects elicited by adult wormsand which are beneficial for the survival of the offspring[15]. It is likely that these protective effects persisted inIL-4 KO or BALB/c IL-4 KO and BALB/c IL-4Ra KOmice even after the elimination of adult worms. Tofurther show that the adult parasite is crucial for theviability of MF, purified MF were injected intraperi-toneally into naive mice. MF killing in BALB/c IL-4KO, BALB/c IL-4Ra KO and BALB/c mice wasequivalent while microfilaraemia in IL-5 KO mice wasfive-fold higher (Table 3) but did not reach levels whichwere observed during the natural infection (Fig. 1A,Table 1). The peritoneal cavity is a natural site to MFinsofar as adult worms can reside there and give birth toMF. It appears that the high microfilaraemia duringnatural infection is associated with the presence of adultworms which were shown in this model to elicit immu-nosuppressive effects beneficial for the survival of MF.Interestingly, this effect required IL-10, and this arguesfor an opposing role of this cytokine compared to IL-4and IL-5 in parasite containment [15].
This study shows that IL-4 and IL-5 lead to thecontainment of different parasite developmental stages.While IL-5 is essential for the control of all develop-mental stages, IL-4 is involved in the containment ofMF. IL-4 acts via IL-4Ra and not directly on the worm.There is no evidence as to a role of IL-13 in addition toIL-4 in parasite containment.
Acknowledgements We are indebted to T. Schueler and staff fortheir help with the maintenance of the parasite life cycle, and to B.Richter for animal genotyping and H. Mossmann for help withmaintaining the BALB/c IL-4 KO and BALB/c IL-4Ra KO mice.The work formed part of a doctoral study by L. Volkmann at theFaculty of Biology, University of Hamburg, Germany. This studyreceived financial support from the Deutsche Forschungsgemeins-chaft (grant Ho 2009/1-3 to A. Hoerauf).
References
1. Al-Qaoud KM, Fleischer B, Hoerauf A (1998) The Xid defectimparts susceptibility to experimental murine filariosis – asso-ciation with a lack of antibody and IL-10 production by B cellsin response to phosphorylcholine. Int Immunol 10:17–25
2. Al-Qaoud KM, Pearlman E, Klukowski J, Hartung T, Flei-scher B, Hoerauf A (2000) A new mechanism for IL-5 depen-dent helminth control: neutrophil accumulation andneutrophil-mediated worm encapsulation in murine filariasisare abolished in the absence of IL-5. Int Immunol 12:899–908
3. Al-Qaoud KM, Taubert A, Zahner H, Fleischer B, Hoerauf A(1997) Infection of BALB/c mice with the filarial nematodeLitomosoides sigmodontis: role of CD4+ T cells in controllinglarval development. Infect Immun 65:2457–2461
4. Bain O, Wanji S, Vuong PN, Marechal P, Le Goff L, Petit G(1994) Larval biology of six filariae Onchocercinae in a verte-brate host. Parasite 1:241–254
5. Bancroft AJ, Grencis RK, Else KJ, Devaney E (1993) Cytokineproduction in BALB/c mice immunized with radiation atten-uated third stage larvae of the filarial nematode, Brugia pah-angi. J Immunol 150:1395–1402
6. Brattig NW, Buttner DW, Hoerauf A (2001) Neutrophilaccumulation around Onchocerca worms and chemotaxis ofneutrophils are dependent on Wolbachia endobacteria.Microbes Infect 3:439–446
7. Brombacher F (2000) The role of interleukin-13 in infectiousdiseases and allergy. Bioessays 22:646–656
8. Chandrashekar R, Rao RU, Rajasekariaf RG, SubrahmanyamD (1984) Separation of viable microfilariae free of blood cellson Percol gradient. J Helminthol 58:69–70
9. Finkelman FD, Pearce EJ, Urban JFJ, Sher A (1991) Regu-lation and biological function of helminth-induced cytokineresponses. Immunol Today 12:62–66
10. Folkard SG, Hogarth PJ, Taylor MJ, Bianco AE (1996) Eo-sinophils are the major effector cells of immunity to microfil-ariae in a mouse model of onchocerciasis. Parasitology112:323–329
11. Hilton DJ, Zhang JG, Metcalf D, Alexander WS, Nicola NA,Willson TA (1996) Cloning and characterization of a bindingsubunit of the IL-13 that is also a component of the IL-4receptor. Proc Natl Acad Sci USA 93:497–501
12. Hoerauf A, Fleischer B (1997) Immune responses to filarialinfection in laboratory mice. Med Microbiol Immunol185:207–215
13. Hoerauf A, Volkmann L, Hamelmann C, Adjei O, AutenriethIB, Fleischer B, Buttner DW (2000) Endosymbiotic bacteria inworms as targets for a novel chemotherapy in filariasis. Lancet355:1242–1243
14. Hoffmann KF, James SL, Cheever AW, Wynn TA (1999)Studies with double cytokine-deficient mice reveal that highlypolarized Th1- and Th2-type cytokine and antibody responsescontribute equally to vaccine-induced immunity to Schistosomamansoni. J Immunol 163:927–938
15. Hoffmann WH, Pfaff AW, Schulz-Key H, Soboslav PT (2001)Determinants for resistance and susceptibility to microfilare-mia in Litomosoides sigmodontis filariasis. Parasitology122:641–649
16. Jankovic D, Kullberg MC, Noben-Trauth N, Cacpar JM,Ward JM, Cheever AW, Pal WE, Sher A (1999) Schistosome-infected IL-4 receptor knockout mice, in contrast to IL-4 KOmice, fail to develop granulomatous pathology while main-taining the same lymphokine expression profile. J Immunol163:337–342
17. Johnson EH, Schynder-Candrian S, Rajan TV, Nelson FK,Lustigman S, Abraham D (1998) Immune responses to thirdstage larvae of Onchocerca volvulus in interferon-gamma andinterleukin-4 knockout mice. Parasite Immunol 20:319–324
18. King CL, Malhotra I, Jia X (1996) Schistosoma mansoni:protective immunity in IL-4-deficient mice. Exp Parasitol84:245–252
30

19. Kopf M, Brombacher F, Hodgkin PD, Ramsay AJ, MilbourneEA, Dai WJ, Ovington KS, Behm CA, Kohler G, Young IG,Matthaei KI (1996) IL-5-deficient mice have a developmentaldefect in CD5+ B-1 cells and lack eosinophilia but havenormal antibody and cytotoxic T cell responses. Immunity4:15–24
20. Lange AM, Yutanawiboonchai W, Scott P, Abraham D (1994)IL-4 and IL-5-dependent protective immunity to Onchocercavolvulus infective larvae in BALB/cBYJ mice. J Immunol153:205–211
21. Lawrence RA (1996) Lymphatic filariasis: what mice can tellus. Parasitol Today 12:267–271
22. Le Goff L, Marechal P, Petit G, Taylor DW, Hoffmann W,Bain O (1997) Early reduction of the challenge recovery ratefollowing immunization with irradiated infective larvae in afilaria mouse system. Trop Med Int Health 2:1170–1174
23. Le Goff L, Loke PN, Ali HF, Taylor DW, Allen JE (2000) IL-5is essential for vaccine-mediated immunity but not innate re-sistance to a filarial parasite. Infect Immun 68:2513–2517
24. Marechal P, Goff L, Petit G, Diagne M, Taylor DW, Bain O(1996) The fate of the filaria Litomosoides sigmodontis in sus-ceptible and naturally resistant mice. Parasite 3:25–31
25. Martin C, Al-Qaoud KM, Bain O, Paehle K, Fleischer B,Hoerauf A (2000) IL-5 is essential for protection afterimmunization against murine filariasis but not during primaryinfection. Med Microbiol Immunol (Berl) 189:67–74
26. Martin C, Le Goff L, Ungeheuer MN, Vuong PN, Bain O(2000) Drastic reduction of a filarial infection in eosinophilicIL-5 transgenic mice. Infect Immun 68:3651–3656
27. Martin C, Saeftel M, Vuong PN, Babayan S, Fischer K, BainO, Hoerauf A (2001) B-cell deficiency suppresses vaccine-in-duced protection against murine filariasis but does not increasethe recovery rate for primary infection. Infect Immun 69:7067–7073
28. Mohrs M, Ledermann B, Kohler G, Dorfmuller A, Gessner A,Brombacher F (1999) Differences between IL-4- and IL-4 re-ceptor alpha-deficient mice in chronic leishmaniasis reveal a
protective role for IL-13 receptor signaling. J Immunol162:7302–7308
29. Mountford AP, Hogg KG, Coulson PS, Brombacher F (2001)Signaling via IL-4 receptor alpha-chain is required for suc-cessful vaccination against schistosomiasis in BALB/c mice.Infect Immun 69:228–236
30. Nelms K, Keegan AD, Zamorano J, Ryan J, Paul WE (1999)The IL-4 receptor: signaling mechanisms and biologic func-tions. Annu Rev Immunol 17:701–738
31. Noben-Trauth N, Kohler G, Burki K, Ledermann B (1996)Efficient targeting of the IL-4 gene in a BALB/c embryonicstem cell line. Transgenic Res 5:487–497
32. Ogbogu VC, Storey DM (1988) In vivo encapsulation andkilling of Litomosoides carinii in white rats. Parasite Immunol10:219–235
33. Petit G, Diagne M, Marechal P, Owen D, Taylor D, Bain O(1992) Maturation of the filaria Litomosoides sigmodontis inBALB/c mice; comparative susceptibility of nine other inbredstrains. Ann Parasitol Hum Comp 67:144–150
34. Saeftel M, Volkmann L, Korten S, Brattig N, Al-Qaoud KM,Fleischer B, Hoerauf A (2001) Lack of IFN-c confers impairedneutrophil granulocyte function and imparts prolonged sur-vival of adult filarial worms in murine filariasis. MicrobesInfect 69:2252–2259
35. Volkmann L, Saeftel M, Bain O, Fischer K, Fleischer B,Hoerauf A (2001) IL-4 is essential for the control of microfil-ariae in murine infection with the filaria Litomosoides sigm-odontis. Infect Immun 69:2950–2956
36. Yates JA, Higashi GI (1985) Brugia malayi: vaccination ofjirds with 60cobalt-attenuated infective stage larvae protectsagainst homologous challenge. Am J Trop Med Hyg 34:1132–1137
37. Zurawski G, de Vries JE (1994) IL-13, an IL-4-like cytokinethat acts on monocytes and B cells, but not on T cells.Immunol Today 15:19–26
31





























