Embed Size (px)
Citation preview

Reduktion der Prädominanz unter binokularer Rivalität
durch Inversionseffekt und Augenwechsel
Bachelorarbeit
Johannes Gutenberg-Universität Mainz
Fachbereich 02 – Sozialwissenschaften, Medien und Sport
Psychologisches Institut, Abteilung Methodenlehre und Statistik
vorgelegt
von
Martin Alexander Brehmen
geb. am 28. Mai 1982
Matrikelnummer: 2678456
Mainz, 07. August 2014
1. Gutachter: Prof. Dr. Günter Meinhardt
2. Gutachter: Dr. Malte Persike

Inversionseffekt und Augenwechsel reduzieren Prädominanz unter binokularer Rivalität
2
Zusammenfassung Binokulare Rivalität entsteht, wenn zwei sehr unterschiedliche Bilder jeweils einem
Auge präsentiert werden. Statt zur Verschmelzung der Bilder kommt es zur wechsel-
haften Wahrnehmung der Einzelbilder. In dieser Studie wurden diesbezüglich zwei
Experimente an zwölf Studierenden durchgeführt. Der menschlichen Bildverarbeitung
von Gesichtern wird eine besondere Rolle attestiert, insbesondere dann, wenn diese in
aufrechter Orientierung präsentiert werden. So zeigten sich in der Vergangenheit
Vorteile gegenüber invertierten Gesichtern als auch gegenüber anderen Objekten wie
Häusern. Im ersten Experiment wurde unter binokularer Rivalität der Inversionseffekt
von Gesichtern überprüft und mit dem von Häusern verglichen. Beide Stimuli erreichten
deutlich längere Dominanzzeiten in der aufrechten Orientierung mit leichten Vorteilen
für Gesichter. Im zweiten Experiment wiesen Gesichter im direkten Vergleich mit
Häusern längere Dominanzzeiten auf, sowohl in aufrechter als auch in invertierter
Orientierung, was für einen generellen Verarbeitungsvorteil von Gesichtern spricht.
Gemessen an den Verarbeitungshypothesen zu binokularer Rivalität sprechen diese
Ergebnisse für eine späte, sprich stimulusbezogene neuronale Verarbeitung. Um
diesbezüglich noch weitere Erkenntnisse zu sammeln, waren in beiden Experimenten
zwei zusätzliche Bedingungen integriert. Zum einen ein Wechsel des dominanten
Stimulus auf das kontralaterale Auge, zum anderen ein gleichzeitiges Aufblinken der
Stimuli. Beide Bedingungen offenbarten starke Einbußen der Dominanzzeiten. Mögliche
Schlussfolgerungen bezüglich der augen- oder stimulusbezogenen Verarbeitung werden
diskutiert.
Schlüsselwörter: binokulare Rivalität; Inversionseffekt; Gesichtsverarbeitung; Augenwechsel

Inversionseffekt und Augenwechsel reduzieren Prädominanz unter binokularer Rivalität
3
Abstract
Binocular rivalry occurs when one eye is presented with an image which differs greatly
from the one which is presented to the other eye. Rather than the two images being
combined into one, they are perceived as two separate, alternating images. As part of
this study, two experiments were conducted on twelve students. Human face-specific
processing was found to play a special role, especially when images of upright faces are
presented. It has been shown in the past that these take precedence over inverted faces
as well as over other objects, such as houses. The first experiment tested the face inver-
sion effect under binocular rivalry conditions, and compared it to the inversion effect
experienced when images of houses are used. Both stimuli achieved much longer
durations of dominance epochs when upright images were presented; the effect was
slightly more pronounced when using faces. The second experiment showed images of
faces – both upright and inverted – to be highly dominant when directly compared to
images of houses. Considering hypotheses on how binocular rivalry may be processed,
these results indicate a late, i.e. stimulus-driven level of processing. In order to gather
more data on this, both experiments included two additional conditions: for one thing,
the images being presented to the dominant and suppressed eye are interchanged, and
for another thing, both stimuli were flashed simultaneously. Both conditions resulted in
significantly shorter durations of dominance epochs. The possible implications of these
findings on the level of processing are being discussed.
Keywords: binocular rivalry; inversion effect; face-specific processing; eye-reversal

Inversionseffekt und Augenwechsel reduzieren Prädominanz unter binokularer Rivalität
4
Inhaltsverzeichnis
1 Einleitung……………………………………………………………………………………………………………..5
2 Methodik…………………………………………………………………………………………………………….10
2.1 Untersuchungsdesign……………………………………………………………………...…………….10
2.2 Versuchspersonen………………………………………………………………………………………..10
2.3 Stimuli………………………………………………………………………………………………………….11
2.4 Instrumente………………………………………………………………………………………………….11
2.5 Aufbau………………………………………………………………………………………………………….12
3 Ergebnisse…………………………………………………………………………………………………………..13
3.1 Experiment I (Inversionseffekte)……………………………………………………………………13
3.2 Experiment II (Stimulustyp)…………………………………………………………………………...16
3.3 Switcheffekte…………………………………………………………………………………………………18
4 Diskussion…………………………………………………………………………………………………………..21
5 Anhang…………………………………………………………………………………………………………........24
6 Literaturverzeichnis……………………………………………………………………………………………25

Inversionseffekt und Augenwechsel reduzieren Prädominanz unter binokularer Rivalität
5
1 Einleitung
Warum besitzen wir zwei Augen? Der bedeutendste Vorteil liegt sicherlich im stereo-
skopischen Sehen, dem Sehen mit beiden Augen, mit dessen Hilfe wir einen räumlichen
Tiefeneindruck bei nahen Gegenständen gewinnen können. Dieser Eindruck entsteht
durch binokulare Fusion, d.h. das Gehirn verrechnet die Informationen, die ihm beide
Augen liefern, und lässt diese zu einem einheitlichen Bild verschmelzen (Birbaumer &
Schmidt, 2010). Präsentiert man jedoch dem linken und dem rechten Auge zwei sehr
unterschiedliche Bilder, schafft es das Gehirn nicht, diese zu einem beständigen und
schlüssigen Gesamtbild zu vereinen. Stattdessen erlebt man eine wechselnde Wahr-
nehmung der Stimuli. Dieses visuelle Phänomen heißt binokulare Rivalität.
Die erste systematische Untersuchung im Bereich der binokularen Rivalität veröffent-
lichte der britische Physiker Charles Wheatstone im Jahre 1838. Er begann mit seinen
Forschungsarbeiten in Folge seiner Erfindung des Stereoskops, mit dem jedem Auge ein
unterschiedliches Bild präsentiert werden kann (Blake & Logothetis, 2002). In anderen
sensorischen Bereichen konnten sehr ähnliche Beobachtungen gemacht werden. So
zeigten sich rivalisierende Sinneseindrücke sowohl beim Hören (Deutsch, 1974) als
auch beim Geruchs- (Zhou & Chen, 2009) und Tastsinn (Carter, Konkle, Wang, Hayward
& Moore, 2008). Dies lässt vermuten, dass das Gehirn eine allgemeine Form der
Verarbeitung mehrdeutiger Wahrnehmungsinformationen besitzt.
Bei der binokularen Rivalität konkurrieren optische Stimuli um die visuelle Wahrneh-
mung. Ist einer der beiden dominant, d.h. deutlich erkennbar, ist der andere während
dieser Zeit supprimiert und kann zumindest nicht bewusst gesehen werden. Die Wahr-
nehmung der Bilder wechselt unwillkürlich (Wheatstone, 1838) und lässt sich nur in
wenigen Fällen bewusst steuern, wie z.B. von tibetanischen Mönchen (Carter et al.,
2005), die mittels Meditation eine fokussiertere Aufmerksamkeit erreichen können.
Beim Wechsel der Dominanz wird dabei häufig eine wellenartige Ausbreitung beob-
achtet (Wilson, Blake & Lee, 2001). Die Dominanzphasen dauern interindividuell unter-
schiedlich lange an und wechseln unregelmäßig (Fox & Herrmann, 1967). Ihre Dauer
hängt von zahlreichen Faktoren ab, auf die im Folgenden noch näher eingegangen wird.
Die Häufigkeitsverteilung der Dauer der Dominanzphasen entspricht verglichen an
empirischen Daten am ehesten einer Beta- und einer Gammaverteilung (Brascamp, van
Ee, Pestman & van den Berg, 2005).

Inversionseffekt und Augenwechsel reduzieren Prädominanz unter binokularer Rivalität
6
Da sich nicht die Stimuli, jedoch deren bewusste Wahrnehmung verändern, leistet die
binokulare Rivalität einen wichtigen Beitrag zur Erforschung des Bewusstseins. Die
zentrale Fragestellung dabei ist, wie das Gehirn zu einer bewussten Erfahrung gelangt
und auf welcher Stufe der Verarbeitung sich eine Lösung des Wettstreits darbietet. Eine
frühe Verarbeitung spräche hierbei für eine Rivalität der Augen, eine späte für die der
Stimuli. Lange Zeit galt die Vermutung, binokulare Rivalität sei eine gegenseitige Inhi-
bition zwischen den monokularen Neuronen im primären visuellen Kortex (V1) und es
fände eine frühe Verarbeitung statt (Blake, 1989), d.h. das Bild jedes einzelnen Auges
würde abwechselnd unterdrückt.
Unterstützende Evidenz findet sich in einer Studie, in der sich mit Hilfe eines fMRT
(funktionelle Magnetresonanztomographie) zeigte, dass die neuronale Aktivität in
monokularen Regionen von V1 mit der jeweiligen Dominanzwahrnehmung wechselte
(Tong & Engel, 2001). Auch sprechen die Ergebnisse von Blake, Westendorf und Overton
(1980) für eine Rivalität der Augen. In ihrem Experiment ließen sie die Versuchs-
personen den jeweils dominanten Stimulus per Knopfdruck mitteilen und tauschten in
unregelmäßigen Abständen die Stimuli auf das jeweils kontralaterale Auge mit dem
Ergebnis, dass der zuvor unterdrückte Stimulus nun sehr viel häufiger Dominanz er-
langte. Dieses Ergebnis führt die Autoren zu dem Schluss, dass die dominante Wahr-
nehmung vom Auge abhängig ist und nicht vom Stimulus und dessen Bedeutung.
Für die Rivalität der Augen spricht ebenfalls, dass früh verarbeitete Eigenschaften eines
Stimulus entscheidenden Einfluss auf seine Prädominanz haben. Als Prädominanz
bezeichnet man das Verhältnis an der Gesamtzeit, die ein Stimulus unter binokularer
Rivalität zu sehen ist. Verändert man einige seiner allgemeinen Bildeigenschaften,
ändert sich auch seine "Stärke", d.h. die Dominanzzeitverhältnisse verschieben sich
(Levelt, 1968). Ein kontrastreiches Bild ist "stärker" als ein kontrastarmes, d.h. es
erreicht vergleichsweise längere Dominanzzeiten (Mueller & Blake, 1989). Ähnlich
verhält es sich mit der Helligkeit. Erhöht sie sich bei einem Stimulus, so steigt dessen
Dominanzzeit an. Sie fällt allerdings bei einem sehr hohen Helligkeitsgrad wieder ab
(Kaplan & Metlay, 1964). Dieser Funktionsverlauf, der in etwa einem invertierten U
entspricht, findet sich ebenfalls bei der Ortsfrequenz eines Stimulus wieder (OShea, Sims
& Govan, 1997). Mittlere Ortsfrequenzen sorgen demnach für die größten Dominanz-
zeiten.

Inversionseffekt und Augenwechsel reduzieren Prädominanz unter binokularer Rivalität
7
Eine ganze Reihe von Studienergebnissen stellte die Theorie, binokulare Rivalität be-
gründe sich ausschließlich mit der Rivalität der Augen, in Frage. Schon Bagby (1957)
zeigte mit seinem Experiment, dass die Familiarität eines Stimulus dessen Dominanz-
zeiten beeinflusst. Dies erreichte Bagby, indem er einer Gruppe von Mexikanern und
einer Gruppe von Amerikanern rivalisierende Stimuli präsentierte, von denen jeweils
einer der mexikanischen und der andere der amerikanischen Kultur zugehörig war. Die
Ergebnisse wiesen auf eine höhere Prädominanz der vertrauten Stimuli hin.
Ein aktuellerer Versuch zeigte, dass die Imagination eines Stimulus dessen Dominanz-
zeit im nachfolgenden Experiment erhöht (Pearson, Clifford & Tong, 2008). Zudem
bestärken neurophysiologische Ergebnisse die Annahme, dass der Stimulus entschei-
denden Einfluss auf die Prädominanz hat. So fanden Leopold und Logothetis (1996) eine
hohe Korrelation zwischen der neuronalen Aktivität in höheren kortikalen Ebenen (V4)
und der Dominanz eines Stimulus. Auch im inferotemporalen Kortex konnten Veränder-
ungen neuronaler Aktivität im Zusammenhang mit dem wahrgenommenen Stimulus
nachgewiesen werden (Sheinberg & Logothetis, 1997). Noch klarer wird dies, wenn man
das Experiment von Kovacs, Papathomas, Yang und Feher (1996) betrachtet. Die For-
scher verwendeten als Stimuluspaar ein Bild von einem Affengesicht und ein Bild von
einem Waldstück. Die beiden Bilder legten sie und flickten sie anschließend so zu-
sammen, dass sie jeweils Inhalte vom anderen Bild an den gleichen Stellen der Ur-
sprungsbilder aufwiesen. Unter binokularer Rivalität kam es bei den Versuchspersonen
dennoch zur wechselweisen Wahrnehmung der beiden ursprünglichen Stimuli. Dies ließ
auf eine neuronale Verarbeitung schließen, welche die zusammengeführten Infor-
mationen beider Augen nutzt und sprach daher gegen die Theorie der Rivalität der
Augen.
Dies deckt sich mit Ergebnissen einer Studie von Tong, Nakayama, Vaughan und Kan-
wisher (1998), die eine enge Verknüpfung zwischen visuellem Bewusstsein und neuro-
naler Aktivität in höheren Hirnzentren aufzeigen konnten. Die Versuchspersonen er-
lebten dabei die binokulare Rivalität eines Gesichts- und eines Hausstimulus. Dabei
wechselte mit der berichteten Wahrnehmung auch die Aktivität in Hirnregionen, die mit
der spezifischen Objektverarbeitung assoziiert ist: Bei Dominanz des Gesichts im fusi-
formen Gesichtsareal (Kanwisher & Yovel, 2006), bei Dominanz des Hauses mit dem

Inversionseffekt und Augenwechsel reduzieren Prädominanz unter binokularer Rivalität
8
parahippokampalen Ortsareal (parahippocampal place area, PPA; Epstein & Kanwisher,
1998).
Letztlich ist bis heute keine klare Entscheidung für eine der beiden Theorien möglich, da
beide im Stande sind, große Teile der gesammelten Erkenntnisse bezüglich binokularer
Rivalität zu erklären. Daher geht die Forschung mittlerweile zunehmend davon aus, dass
es sich um ein Hybridmodell handelt, das beide Ansätze integriert (Tong, Meng & Blake,
2006). Demnach wirken sowohl frühe als auch späte neuronale Verarbeitungsmecha-
nismen zusammen.
Eine beachtliche Studie, in der es um den Einfluss von bedeutungsvollen Inhalten auf
binokulare Rivalität geht, verfassten Yu und Blake (1992). Sie zeigten beispielsweise,
dass ein Gesicht eine größere Prädominanz gegenüber einem unbedeutenden Stimulus
aufweist, auch wenn Bildeigenschaften wie Amplitudenspektrum, Helligkeit und Kon-
trast identisch sind. In einem weiteren Experiment dieser Studie verschleierten die
Forscher den bedeutungsvollen Inhalt eines Stimulus, so dass er für die
Versuchspersonen im ersten Durchgang nicht bewusst wahrnehmbar war. Dennoch
konnte er einen Prädominanzvorteil gegenüber einem zufälligen Punktmuster auf-
weisen, was für einen großen Einfluss der konfiguralen Eigenschaften von Stimuli
spricht. Auch zeigten Yu und Blake, dass die aufrechte Orientierung von bedeutungs-
vollen Stimuli höhere Dominanzzeiten gegenüber ihren Inversen bewirkt. Dies deckt
sich mit den Erkenntnissen von Engel (1956), der zeigte, dass aufrechte Gesichter eine
stärkere Prädominanz erreichen als invertierte Gesichter.
Die Gesichtswahrnehmung soll auch in dieser Studie eine entscheidende Rolle spielen.
Das Erkennen von Gesichtern besitzt eine enorme biologische und soziale Bedeutung.
Daher liegt die Vermutung nahe, dass sich evolutionsgeschichtlich spezielle Mechanis-
men entwickelt haben, die diese Aufgabe per se erleichtern. Andererseits erlangt der
Mensch im Laufe seines Lebens aber auch eine immense Erfahrung im Umgang mit Ge-
sichtern, so dass man sich zu einem wahren Gesichtsexperten entwickelt, was wiederum
zu besonderen Fertigkeiten in der Verarbeitung führen könnte (Goldenberg, 2012).
Ähnlich der Frage nach dem genauen Ort der Verarbeitung von binokularer Rivalität
konnte auch diese Frage bislang nicht vollends geklärt werden.
Die Wiedererkennung von Gesichtern erwies sich in Gedächtnisexperimenten als deut-
lich beeinträchtigt, wenn man diese invertierte (Yin, 1969) mit in der Folge stark redu-
zierter Genauigkeit und erhöhten Reaktionszeiten. Dieses Phänomen, das als "face in-

Inversionseffekt und Augenwechsel reduzieren Prädominanz unter binokularer Rivalität
9
version effect" bezeichnet wird, zeigte sich jedoch nicht bei der Wiedererkennung
anderer monoorientierter Objekte, die für gewöhnlich nur in der aufrechten Orien-
tierung gesehen werden, wie beispielsweise Flugzeuge oder Häuser. Man vermutet
daher, dass Gesichter holistisch, also ganzheitlich, verarbeitet werden.
Im Gegensatz zu anderen Objekten, bei denen die Verarbeitung auf einzelnen Merk-
malen basiert, stehen bei Gesichtern konfigurale Eigenschaften wie die räumliche Orga-
nisation bestimmter Gesichtsareale im Vordergrund, was den Menschen zu einem Ex-
perten der Gesichtserkennung macht (Maurer, Le Grand & Mondloch, 2002).
Dementgegen steht eine alternative Erklärung von Diamond und Carey (1986), deren
Befunde gegen einen speziellen Verarbeitungsmodus für Gesichter sprechen. Sie konn-
ten belegen, dass der Inversionseffekt vor allem dann zu beobachten ist, wenn man über
große Expertise bezüglich einer Objektgattung verfügt. Sie zeigten dies anhand von Hun-
deexperten, die bei binokularer Rivalität mit Bildern ihnen wohlbekannter Hunderassen
ähnlich große Inversionseffekte aufwiesen, wie sie sonst nur für menschliche Gesichter
zu beobachten sind. Eine aktuellere Betrachtung rückt von dieser Sichtweise allerdings
wieder etwas ab und lässt der Gesichtsverarbeitung weiterhin eine besondere Rolle
zukommen (McKone, Kanwisher & Duchaine, 2007).
Dass die aufrechte Orientierung vor allem bei der Gesichtsverarbeitung vorteilhaft zu
sein scheint, lassen die Ergebnisse einer Studie von Bannerman, Milders, Gelder und
Sahraie (2008) vermuten. Sie konnten nachweisen, dass Gesichter unter binokularer
Rivalität nur dann Prädominanzvorteile gegenüber Häusern aufwiesen, wenn sie in
aufrechter Orientierung präsentiert wurden. Eine weitere Erkenntnis dieser Studie war
der Prä-dominanzvorteil emotionaler Gesichter gegenüber neutralen Gesichtern.
Aufgrund der Forschungsliteratur können somit folgende Annahmen bezüglich der
Dominanzzeiten bei binokularer Rivalität gemacht werden.
Aufrechte Gesichter sollten im Vergleich mit invertierten Gesichtern höhere Dominanz-
zeiten erzielen (1). Es wäre ferner zu vermuten, dass der Inversionseffekt bei Gesich-
tern höher ausfällt im Vergleich zu Häusern (2). Im direkten Vergleich von Gesichtern
und Häusern werden längere Dominanzzeiten bei Gesichtern erwartet (3). Der Wechsel
eines Stimulus auf das kontralaterale Auge sollte sich in der Verkürzung seiner Domi-
nanzzeit widerspiegeln (4).

Inversionseffekt und Augenwechsel reduzieren Prädominanz unter binokularer Rivalität
10
2 Methodik
2.1 Untersuchungsdesign:
Im Rahmen dieser Untersuchung wurden zwei Experimente in einem within subject-
Design realisiert, in denen jeweils zwei Stimuli gegeneinander rivalisierten. In Experi-
ment I waren es pro Bedingung entweder ein aufrechtes Gesicht oder ein aufrechtes
Haus mit ihrem invertierten Gegenstück. Hiermit soll geklärt werden, ob sich ein In-
versionseffekt beim Gesicht zeigt und ob dieser größer ist als bei Häusern.
In Experiment II waren es pro Bedingung ein Gesicht und ein Haus, die entweder beide
aufrecht oder beide invertiert waren (siehe Abb.1). Hiermit sollten Effekte der Stimulus-
art (Gesicht gegen Haus) aufgedeckt werden, also ob Gesichter im direkten Vergleich mit
Häusern höhere Dominanzzeiten erzielen. Bei allen Kombinationen traten zudem zu
Beginn einer Dominanzphase mit einer Wahrscheinlichkeit von je 33% entweder ein
Wechsel der Stimuli auf das jeweils kontralaterale Auge (Switch), ein einmaliges, kurzes
Aufblinken beider Stimuli, bei dem lediglich das hintergründige Rauschmuster sichtbar
blieb (Aufblinken) oder keine Veränderung auf (Normal). Dieser Faktor wird im Folgen-
den als Switchbedingung bezeichnet und hatte die Klärung der Frage zum Ziel, ob es sich
um die Rivalität der Augen oder der Stimuli handelt. Damit Wahrnehmungsveränder-
ungen nach dem Eintreten der Varianten Switch oder Aufblinken nicht verschleiert
wurden, gab es eine Art Refraktärzeit, in der ein Tastendruck keine dieser beiden Vari-
anten erneut eintreten ließ. Daher stellte sich im Rahmen der Auswertung heraus, dass
die Variante ohne Veränderung in etwa 54% der Fälle eintrat, die anderen beiden in
etwa 23% der Fälle.
2.2 Versuchspersonen:
Bei den Versuchspersonen handelte es sich um zwölf Studierende, elf davon im Bereich
der Psychologie und einer im Bereich Kommunikationsdesign.
Bei der Betrachtung des Alters reichte die Range von 20 bis 34 Jahren (MW: 24,58; SD:
4,70). Die Verteilung des Geschlechts der Teilnehmenden liegt bei acht weiblichen und
vier männlichen Personen. Die Sehstärke der Versuchspersonen war normal oder korri-
giert.
Die Teilnahme geschah freiwillig und wurde bei den Psychologiestudierenden mit Ver-
suchspersonenstunden vergütet, die im Rahmen des Bachelorstudiengangs der Psycho-
logie zu erwerben sind. Der Kommunikationsdesignstudierende erhielt eine finanzielle

Inversionseffekt und Augenwechsel reduzieren Prädominanz unter binokularer Rivalität
11
Aufwandsentschädigung (fünf € pro Stunde). Die Versuchspersonen wurden darüber
informiert, dass sie das Experiment jederzeit und ohne negative Folgen pausieren oder
ganz abbrechen können. Alle erhobenen Daten wurden anonymisiert und vertraulich
behandelt.
2.3 Stimuli:
Für die Experimente wurden Fotoaufnahmen von Gesichtern und Häusern verwendet.
Insgesamt standen jeweils acht zur Verfügung. Die Gesichter stammen aus der Radboud
Faces Database (Langner et al., 2010) und weisen mitteleuropäisches Aussehen mit neu-
tralem Gesichtsausdruck auf. Die Bilder der Häuser gehen auf eine Internetrecherche
zurück und zeigen jeweils die Frontansicht mit Dachgiebel. Alle Bilder waren in ein
Rauschmuster eingebettet, welches etwa 60% des Gesamtbildes einnahm. Zudem
wurden sie mit Adobe Photoshop standardisiert bezüglich der Größe (500 x 500 Pixel
Bounding Box), der Graustufung (mit einer Helligkeit von 65 cd/m2), dem Michelson
Kontrast (0,92) sowie der Ortsfrequenz. Die Invertierung der Stimuli entstand durch
Drehung um 180°. Die Bilder wurden auf hellgrauem Hintergrund präsentiert. Beispiele
für alle vier Stimuluskombinationen zeigt Abb. 1.
A B
Abbildung 1: Kombinationen der Stimuli in Experiment I (A) und Experiment II (B)
2.4 Instrumente:
Das Experiment fand in einem Testlabor des psychologischen Instituts der Johannes
Gutenberg – Universität in Mainz statt. Realisiert wurde es auf einem Desktop-PC mit

Inversionseffekt und Augenwechsel reduzieren Prädominanz unter binokularer Rivalität
12
der Experimentalsoftware Inquisit (Version 4). Der verwendete Monitor hatte ein 20
Zoll großes TFT Display mit einer Auflösung von 1650 x 1050 Pixeln und einer Bild-
wiederholungsrate von 60 Hz. Die Versuchspersonen schauten durch ein Spiegelstereo-
skop, dessen Abstand zum Bildschirm 60 cm betrug. Die Mitteilung der Dominanzwahr-
nehmung erfolgte über zwei Tasten eines externen Keyboards. Das verwendete Modell
ermöglichte besonders genaue Zeitmessungen. Die Beleuchtung des Labors wurde bei
mittlerer Helligkeit konstant gehalten.
2.5 Ablauf:
Zu Beginn der ersten Sitzung erfolgte einmalig die Überprüfung der Augendominanz.
Vier der zwölf Versuchspersonen wiesen Linksdominanz auf (jeweils zwei pro
Geschlecht). Das Stereoskop wurde vor jeder Sitzung individuell von den Versuchs-
personen in der Höhe justiert, um eine bequeme Sitzhaltung zu erreichen. Zudem wurde
ein Kalibrierungs-modus dazu genutzt, die Spiegel des Stereoskops so einzustellen, dass
sich beim Durchschauen ein waagerechter und ein senkrechter Stimulus, die jeweils
einem Auge präsentiert wurden, in der Mitte überlagerten und so zu einem Kreuz
fusionierten. Somit war im späteren Experiment eine optimale Überlagerung der Stimuli
gewährleistet.
In der ersten Sitzung erfolgte anschließend ein Lerndurchgang, um zu prüfen, ob die
Versuchspersonen die Stimuli den zuvor festgelegten Tasten des Keyboards korrekt
zuordnen können. Hierzu wurde im randomisierten Wechsel beiden Augen ein Gesicht
oder ein Haus präsentiert, so dass es zu einer eindeutige Wahrnehmung kam, die mit
dem Druck auf die entsprechende Taste mitgeteilt wurde. Dieser Durchgang galt als
erfolgreich und berechtigte zur weiteren Teilnahme, wenn mindestens 95% der Ant-
worten richtig waren. Dies war bei allen Versuchspersonen der Fall.
Der Hauptteil der Sitzungen bestand aus den vier Stimuluskombinationen aufrechtes
Gesicht gegen invertiertes Gesicht (1), aufrechtes Haus gegen invertiertes Haus (2), auf-
rechtes Gesicht gegen aufrechtes Haus (3) und invertiertes Gesicht gegen invertiertes
Haus (4). Die Invertierungen der ersten beiden Bedingungen waren die jeweilige Spie-
gelung der aufrechten Stimuli. In jeder der vier Bedingungen wurde beiden Augen ein
unterschiedlicher Stimulus dargeboten, so dass es zu binokularer Rivalität kam. Aufgabe
der Versuchspersonen war es, bei Dominanz eines Stimulus die zugeordnete Taste so
lange zu drücken, bis die Dominanz verschwindet. Kam es bei der Wahrnehmung zu

Inversionseffekt und Augenwechsel reduzieren Prädominanz unter binokularer Rivalität
13
Mischzuständen, die eine klare Dominanz verhinderten, sollten sie keine der beiden
Tasten drücken. Die Kombinationen wurden randomisiert dargeboten, wobei jede Be-
dingung einmal pro Sitzung durchlaufen werden musste. Dies sollte eventuelle Stör-
größen wie Müdigkeit oder Lerneffekte reduzieren. Die Anzahl an Trials pro Bedingung
lag im Mittel bei 248,44 (SD: 72,62). Ein Trial endete mit dem Loslassen der Taste. Dau-
erte eine Bedingung länger als 15 Minuten, wurde sie frühzeitig abgebrochen und zur
nächsten übergegangen.
Die Versuchspersonen nahmen an vier Sitzungen innerhalb eines Monats teil, die jeweils
zwischen 45 und 100 Minuten dauerten.
3 Ergebnisse
Die Auswertung der Daten erfolgte mit Statistica (Version 9). Aus den absoluten Domi-
nanzzeiten der Versuchspersonen ergaben sich mittlere Dominanzzeiten für alle Bedin-
gungskombinationen, die als abhängige Variable dienten. Experiment I und II wurden
dabei getrennt analysiert. Über sie wurde jeweils eine Varianzanalyse mit Messwieder-
holung (rmANOVA) gerechnet. Als Innersubjektfaktoren dienten die Bedingungen Sti-
mulus (Gesicht oder Haus), Orientierung (aufrecht oder invertiert) und Switch (Augen-
wechsel, Aufblinken oder keine Veränderung), woraus sich insgesamt jeweils zwölf
mögliche Kombinationen für jedes Experiment ergaben.
Die durch die Hypothesen zu erwartenden Unterschiede in den Gruppenmittelwerten,
wurden über a-priori-Kontraste auf ihre Signifikanz hin geprüft. Des Weiteren erfolgte
die Berechnung der Effektgröße Cohens d für abhängige Stichproben (dz = d / σd), wobei
d der Mittelwert der Differenzen und σd die Streuung dieser Differenzen ist (Cohen,
1988).
3.1 Experiment I (Inversionseffekte):
Bezüglich der Inversionseffekte wurde zum einen die Hypothese aufgestellt, dass auf-
rechte Gesichter höhere Dominanzzeiten im Vergleich zu invertierten Gesichtern er-
zielen. Die Ergebnisse, die in Tab. 1 (Seite 14) aufgeführt sind, zeigen ein signifikantes
Ergebnis des Haupteffektes Orientierung (F(1,11) = 26,99, p < 0,001). Der Blick auf die
Mittelwerte lässt vermuten, dass aufrechte Stimuli längere Dominanzzeiten erreichten
im Vergleich mit invertierten ( aufrecht = 2,66s; invertiert = 1,98s).
Inversionseffekt und Augenwechsel reduzieren Prädominanz unter binokularer Rivalität
14
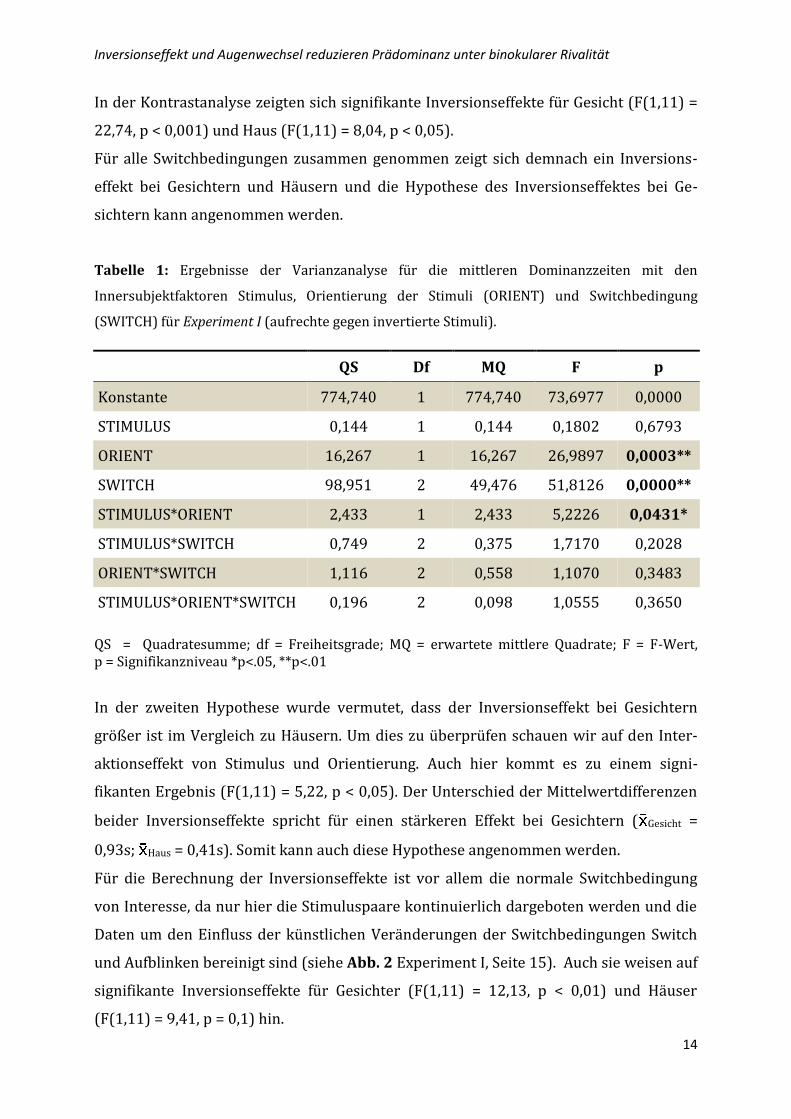
In der Kontrastanalyse zeigten sich signifikante Inversionseffekte für Gesicht (F(1,11) =
22,74, p < 0,001) und Haus (F(1,11) = 8,04, p < 0,05).
Für alle Switchbedingungen zusammen genommen zeigt sich demnach ein Inversions-
effekt bei Gesichtern und Häusern und die Hypothese des Inversionseffektes bei Ge-
sichtern kann angenommen werden.
Tabelle 1: Ergebnisse der Varianzanalyse für die mittleren Dominanzzeiten mit den
Innersubjektfaktoren Stimulus, Orientierung der Stimuli (ORIENT) und Switchbedingung
(SWITCH) für Experiment I (aufrechte gegen invertierte Stimuli).
QS Df MQ F p
Konstante 774,740 1 774,740 73,6977 0,0000
STIMULUS 0,144 1 0,144 0,1802 0,6793
ORIENT 16,267 1 16,267 26,9897 0,0003**
SWITCH 98,951 2 49,476 51,8126 0,0000**
STIMULUS*ORIENT 2,433 1 2,433 5,2226 0,0431*
STIMULUS*SWITCH 0,749 2 0,375 1,7170 0,2028
ORIENT*SWITCH 1,116 2 0,558 1,1070 0,3483
STIMULUS*ORIENT*SWITCH 0,196 2 0,098 1,0555 0,3650
QS = Quadratesumme; df = Freiheitsgrade; MQ = erwartete mittlere Quadrate; F = F-Wert, p = Signifikanzniveau *p<.05, **p<.01
In der zweiten Hypothese wurde vermutet, dass der Inversionseffekt bei Gesichtern
größer ist im Vergleich zu Häusern. Um dies zu überprüfen schauen wir auf den Inter-
aktionseffekt von Stimulus und Orientierung. Auch hier kommt es zu einem signi-
fikanten Ergebnis (F(1,11) = 5,22, p < 0,05). Der Unterschied der Mittelwertdifferenzen
beider Inversionseffekte spricht für einen stärkeren Effekt bei Gesichtern ( Gesicht =
0,93s; Haus = 0,41s). Somit kann auch diese Hypothese angenommen werden.
Für die Berechnung der Inversionseffekte ist vor allem die normale Switchbedingung
von Interesse, da nur hier die Stimuluspaare kontinuierlich dargeboten werden und die
Daten um den Einfluss der künstlichen Veränderungen der Switchbedingungen Switch
und Aufblinken bereinigt sind (siehe Abb. 2 Experiment I, Seite 15). Auch sie weisen auf
signifikante Inversionseffekte für Gesichter (F(1,11) = 12,13, p < 0,01) und Häuser
(F(1,11) = 9,41, p = 0,1) hin.
Inversionseffekt und Augenwechsel reduzieren Prädominanz unter binokularer Rivalität
15
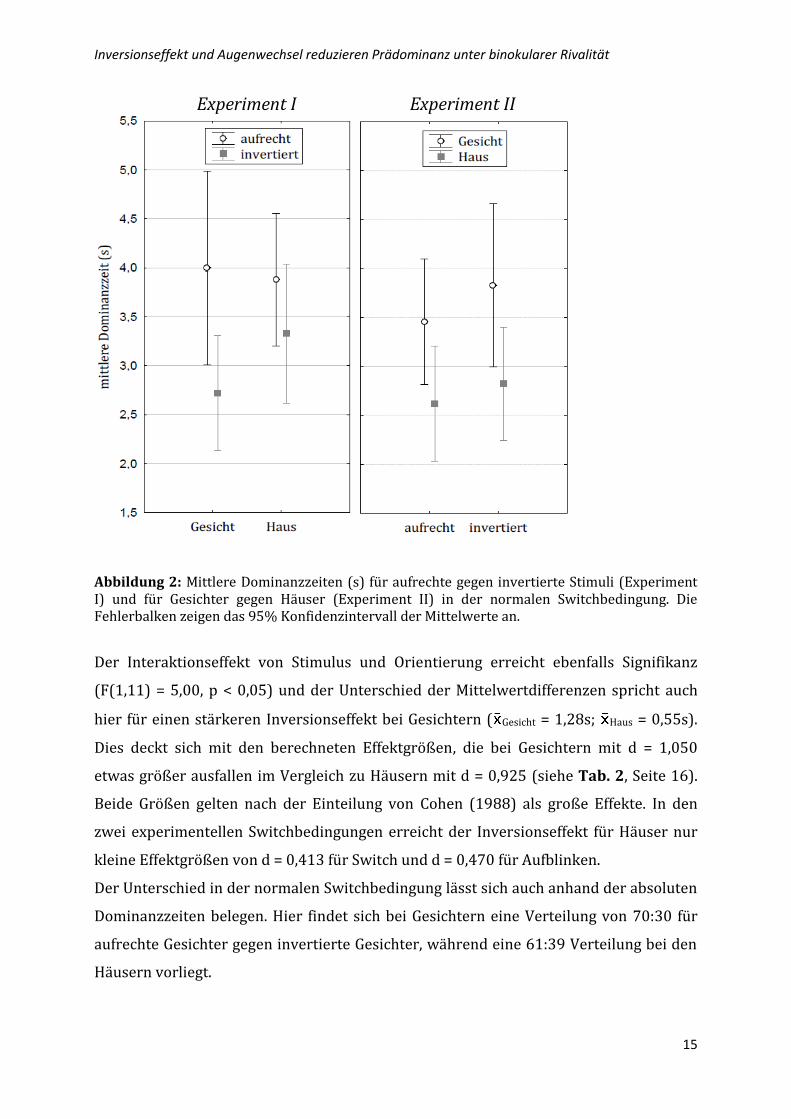
Experiment I Experiment II
Abbildung 2: Mittlere Dominanzzeiten (s) für aufrechte gegen invertierte Stimuli (Experiment I) und für Gesichter gegen Häuser (Experiment II) in der normalen Switchbedingung. Die Fehlerbalken zeigen das 95% Konfidenzintervall der Mittelwerte an.
Der Interaktionseffekt von Stimulus und Orientierung erreicht ebenfalls Signifikanz
(F(1,11) = 5,00, p < 0,05) und der Unterschied der Mittelwertdifferenzen spricht auch
hier für einen stärkeren Inversionseffekt bei Gesichtern ( Gesicht = 1,28s; Haus = 0,55s).
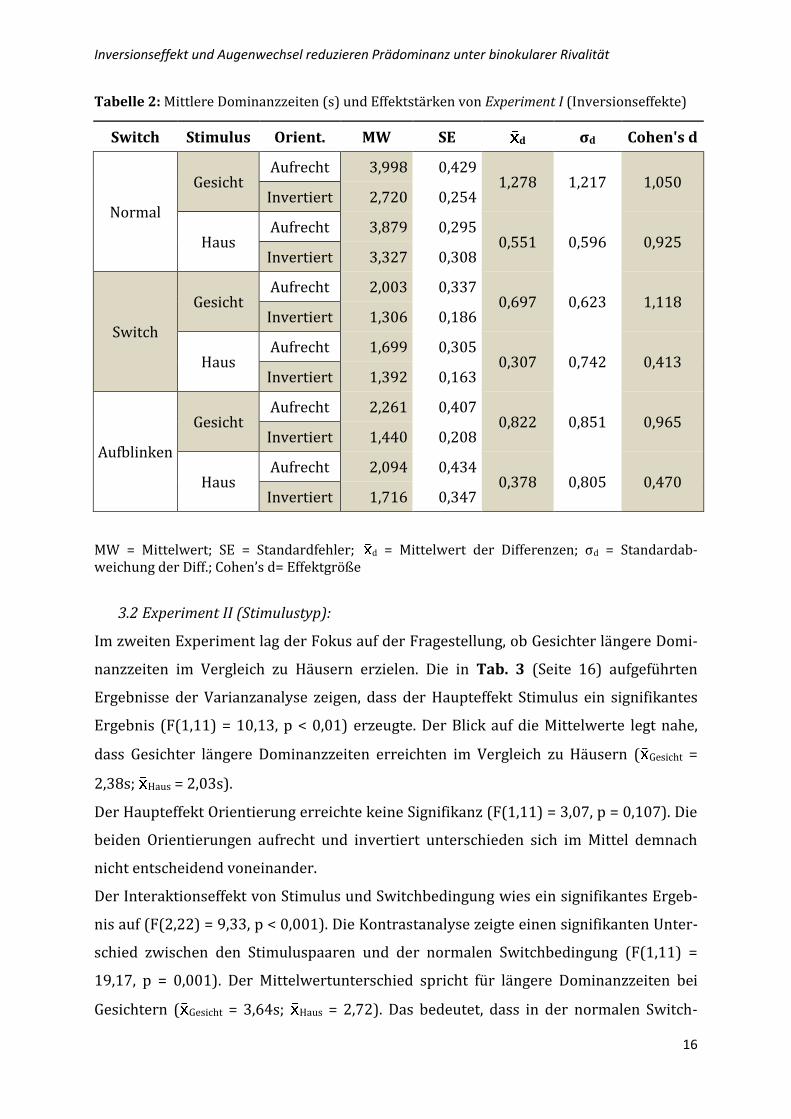
Dies deckt sich mit den berechneten Effektgrößen, die bei Gesichtern mit d = 1,050
etwas größer ausfallen im Vergleich zu Häusern mit d = 0,925 (siehe Tab. 2, Seite 16).
Beide Größen gelten nach der Einteilung von Cohen (1988) als große Effekte. In den
zwei experimentellen Switchbedingungen erreicht der Inversionseffekt für Häuser nur
kleine Effektgrößen von d = 0,413 für Switch und d = 0,470 für Aufblinken.
Der Unterschied in der normalen Switchbedingung lässt sich auch anhand der absoluten
Dominanzzeiten belegen. Hier findet sich bei Gesichtern eine Verteilung von 70:30 für
aufrechte Gesichter gegen invertierte Gesichter, während eine 61:39 Verteilung bei den
Häusern vorliegt.
Inversionseffekt und Augenwechsel reduzieren Prädominanz unter binokularer Rivalität
16
Tabelle 2: Mittlere Dominanzzeiten (s) und Effektstärken von Experiment I (Inversionseffekte)
Switch Stimulus Orient. MW SE d σd Cohen's d
Normal
Gesicht Aufrecht 3,998 0,429
1,278 1,217 1,050 Invertiert 2,720 0,254
Haus Aufrecht 3,879 0,295
0,551 0,596 0,925 Invertiert 3,327 0,308
Switch
Gesicht Aufrecht 2,003 0,337
0,697 0,623 1,118 Invertiert 1,306 0,186
Haus Aufrecht 1,699 0,305
0,307 0,742 0,413 Invertiert 1,392 0,163
Aufblinken
Gesicht Aufrecht 2,261 0,407
0,822 0,851 0,965 Invertiert 1,440 0,208
Haus Aufrecht 2,094 0,434
0,378 0,805 0,470 Invertiert 1,716 0,347
MW = Mittelwert; SE = Standardfehler; d = Mittelwert der Differenzen; σd = Standardab-weichung der Diff.; Cohen’s d= Effektgröße
3.2 Experiment II (Stimulustyp):
Im zweiten Experiment lag der Fokus auf der Fragestellung, ob Gesichter längere Domi-
nanzzeiten im Vergleich zu Häusern erzielen. Die in Tab. 3 (Seite 16) aufgeführten
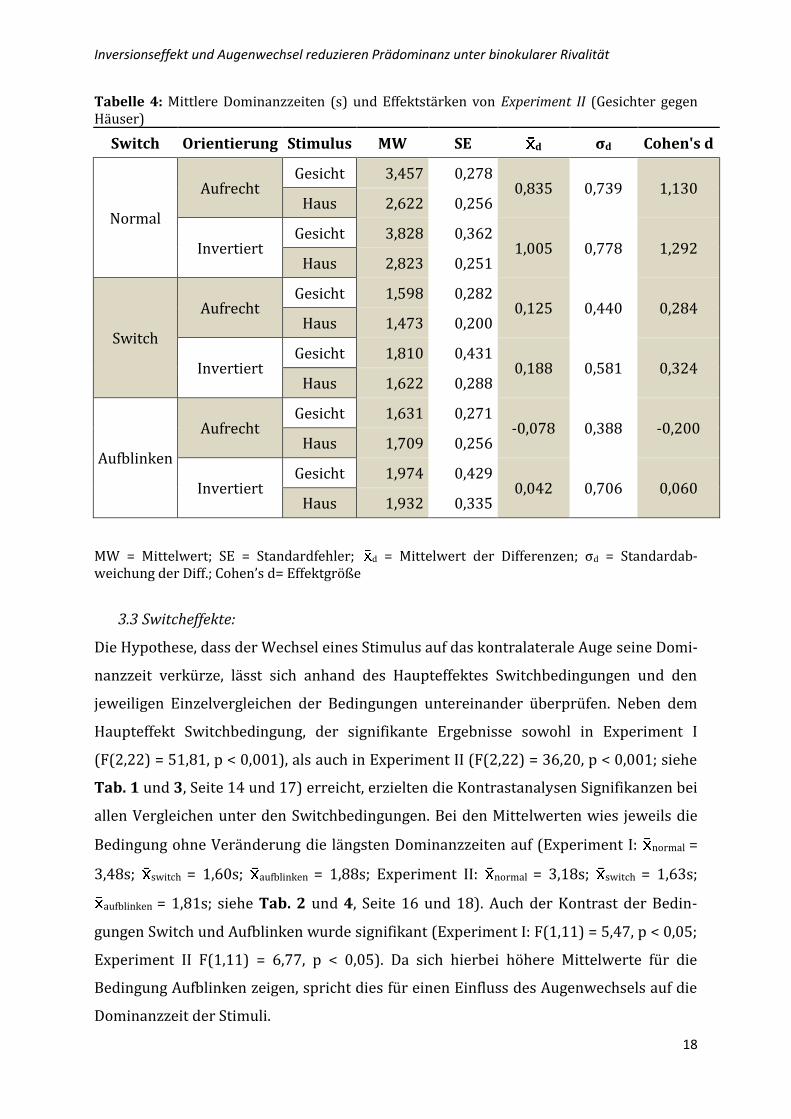
Ergebnisse der Varianzanalyse zeigen, dass der Haupteffekt Stimulus ein signifikantes
Ergebnis (F(1,11) = 10,13, p < 0,01) erzeugte. Der Blick auf die Mittelwerte legt nahe,
dass Gesichter längere Dominanzzeiten erreichten im Vergleich zu Häusern ( Gesicht =
2,38s; Haus = 2,03s).
Der Haupteffekt Orientierung erreichte keine Signifikanz (F(1,11) = 3,07, p = 0,107). Die
beiden Orientierungen aufrecht und invertiert unterschieden sich im Mittel demnach
nicht entscheidend voneinander.
Der Interaktionseffekt von Stimulus und Switchbedingung wies ein signifikantes Ergeb-
nis auf (F(2,22) = 9,33, p < 0,001). Die Kontrastanalyse zeigte einen signifikanten Unter-
schied zwischen den Stimuluspaaren und der normalen Switchbedingung (F(1,11) =
19,17, p = 0,001). Der Mittelwertunterschied spricht für längere Dominanzzeiten bei
Gesichtern ( Gesicht = 3,64s; Haus = 2,72). Das bedeutet, dass in der normalen Switch-

Inversionseffekt und Augenwechsel reduzieren Prädominanz unter binokularer Rivalität
17
bedingung Gesichter signifikant länger wahrgenommen werden als Häuser, wohingegen
in den Switchbedingungen Switch (F(1,11) = 1,07, p = 0,322) und Aufblinken (F(1,11) =
0,01, p = 0,910) dieser Effekt nicht nachgewiesen wurde.
Wie schon bei den Inversionseffekten war auch bei der Berechnung der Effekte des
Stimulustyps vor allem die normale Switchbedingung interessant (siehe Abb. 2 Exper-
iment II, Seite 15). Die um den Einfluss der Switchbedingungen Switch und Aufblinken
bereinigten Daten zeigten ebenfalls signifikante Effekte für aufrechte (F(1,11) = 14,04, p
< 0,01, Gesicht = 3,46s; Haus = 2,62s) und invertierte Orientierung (F(1,11) = 18,37, p =
0,01, Gesicht = 3,83s; Haus = 2,82s).
Die Effektgrößen bestätigen dies: Gesichter erreichten im direkten Vergleich zu Häusern
durchschnittlich längere Dominanzzeiten, sowohl aufrecht mit d = 1,130 als auch inver-
tiert mit d = 1,292 (siehe Tab. 4, Seite 18). Die Effektgrößen sind jeweils als groß einzu-
stufen.
Die Verteilung der absoluten Dominanzzeiten ist mit 58:42 identisch für beide Orien-
tierungen mit jeweils längerer Dominanz von Gesichtern gegenüber Häusern.
Tabelle 3: Ergebnisse der Varianzanalyse für die mittleren Dominanzzeiten mit den Innersubjektfaktoren Stimulus, Orientierung der Stimuli (ORIENT) und Switchbedingung (SWITCH) für Experiment II (Gesichter gegen Häuser).
QS Df MQ F P
Konstante 701,182 1 701,182 62,9003 0,0000
STIMULUS 4,486 1 4,486 10,1310 0,0087**
ORIENT 2,249 1 2,249 3,0731 0,1074
SWITCH 69,375 2 34,688 36,2042 0,0000**
STIMULUS*ORIENT 0,125 1 0,125 1,7417 0,2137
STIMULUS*SWITCH 5,968 2 2,984 9,3264 0,0012**
ORIENT*SWITCH 0,086 2 0,043 0,6462 0,5337
STIMULUS*ORIENT*SWITCH 0,017 2 0,009 0,1453 0,8656
QS = Quadratesumme; df = Freiheitsgrade; MQ = erwartete mittlere Quadrate; F = F-Wert, p = Signifikanzniveau *p<.05, **p<.01
Inversionseffekt und Augenwechsel reduzieren Prädominanz unter binokularer Rivalität
18
Tabelle 4: Mittlere Dominanzzeiten (s) und Effektstärken von Experiment II (Gesichter gegen Häuser)
Switch Orientierung Stimulus MW SE d σd Cohen's d
Normal
Aufrecht Gesicht 3,457 0,278
0,835 0,739 1,130 Haus 2,622 0,256
Invertiert Gesicht 3,828 0,362
1,005 0,778 1,292 Haus 2,823 0,251
Switch
Aufrecht Gesicht 1,598 0,282
0,125 0,440 0,284 Haus 1,473 0,200
Invertiert Gesicht 1,810 0,431
0,188 0,581 0,324 Haus 1,622 0,288
Aufblinken
Aufrecht Gesicht 1,631 0,271
-0,078 0,388 -0,200 Haus 1,709 0,256
Invertiert Gesicht 1,974 0,429
0,042 0,706 0,060 Haus 1,932 0,335
MW = Mittelwert; SE = Standardfehler; d = Mittelwert der Differenzen; σd = Standardab-weichung der Diff.; Cohen’s d= Effektgröße
3.3 Switcheffekte:
Die Hypothese, dass der Wechsel eines Stimulus auf das kontralaterale Auge seine Domi-
nanzzeit verkürze, lässt sich anhand des Haupteffektes Switchbedingungen und den
jeweiligen Einzelvergleichen der Bedingungen untereinander überprüfen. Neben dem
Haupteffekt Switchbedingung, der signifikante Ergebnisse sowohl in Experiment I
(F(2,22) = 51,81, p < 0,001), als auch in Experiment II (F(2,22) = 36,20, p < 0,001; siehe
Tab. 1 und 3, Seite 14 und 17) erreicht, erzielten die Kontrastanalysen Signifikanzen bei
allen Vergleichen unter den Switchbedingungen. Bei den Mittelwerten wies jeweils die
Bedingung ohne Veränderung die längsten Dominanzzeiten auf (Experiment I: normal =
3,48s; switch = 1,60s; aufblinken = 1,88s; Experiment II: normal = 3,18s; switch = 1,63s;
aufblinken = 1,81s; siehe Tab. 2 und 4, Seite 16 und 18). Auch der Kontrast der Bedin-
gungen Switch und Aufblinken wurde signifikant (Experiment I: F(1,11) = 5,47, p < 0,05;
Experiment II F(1,11) = 6,77, p < 0,05). Da sich hierbei höhere Mittelwerte für die
Bedingung Aufblinken zeigen, spricht dies für einen Einfluss des Augenwechsels auf die
Dominanzzeit der Stimuli.
Inversionseffekt und Augenwechsel reduzieren Prädominanz unter binokularer Rivalität
19
Experiment I Experiment II
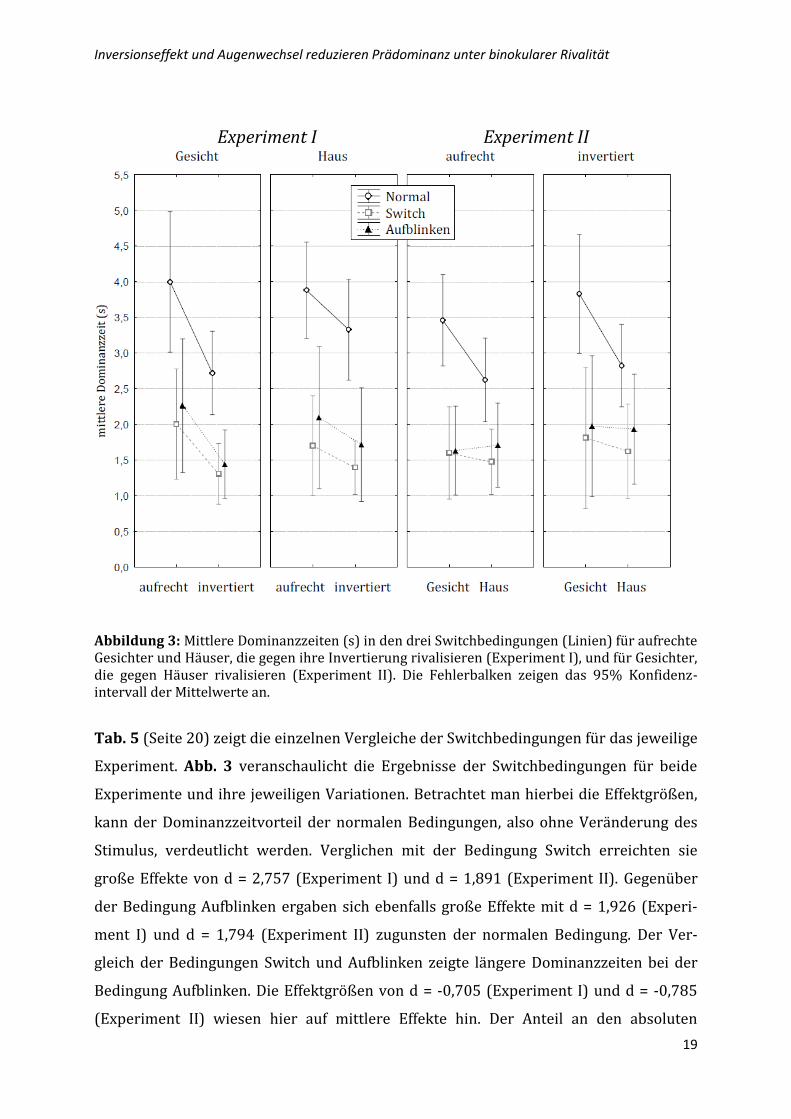
Abbildung 3: Mittlere Dominanzzeiten (s) in den drei Switchbedingungen (Linien) für aufrechte Gesichter und Häuser, die gegen ihre Invertierung rivalisieren (Experiment I), und für Gesichter, die gegen Häuser rivalisieren (Experiment II). Die Fehlerbalken zeigen das 95% Konfidenz-intervall der Mittelwerte an.
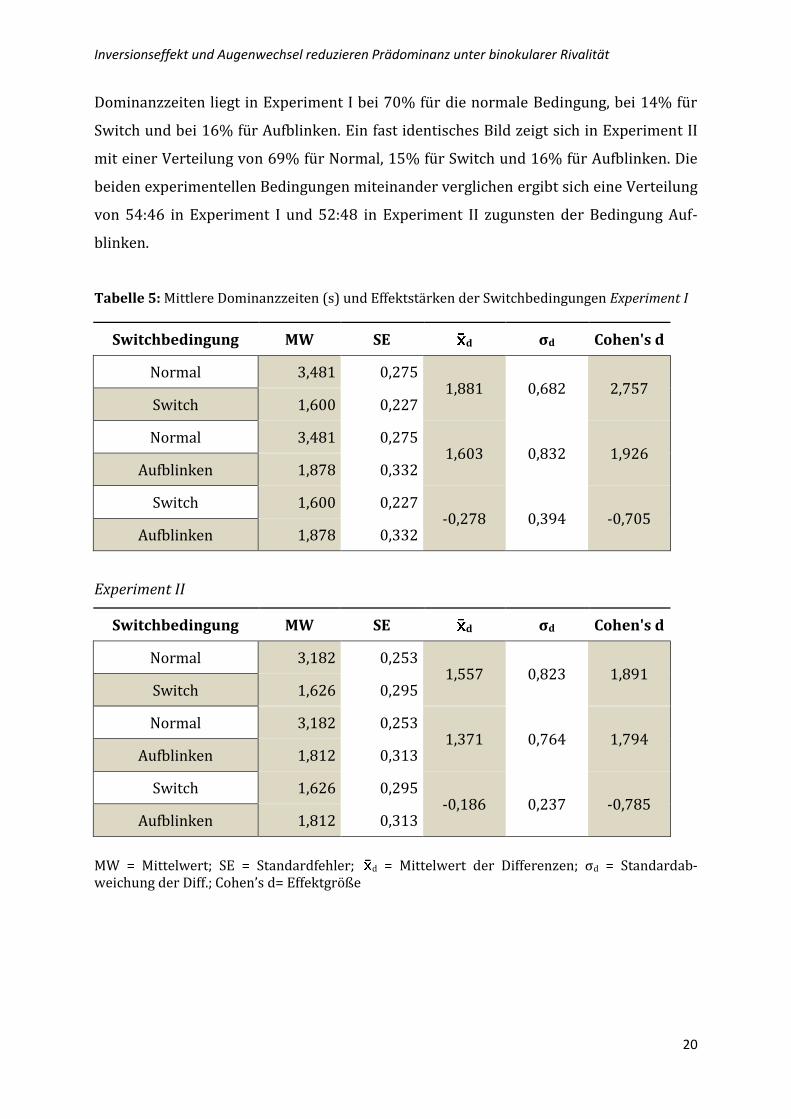
Tab. 5 (Seite 20) zeigt die einzelnen Vergleiche der Switchbedingungen für das jeweilige
Experiment. Abb. 3 veranschaulicht die Ergebnisse der Switchbedingungen für beide
Experimente und ihre jeweiligen Variationen. Betrachtet man hierbei die Effektgrößen,
kann der Dominanzzeitvorteil der normalen Bedingungen, also ohne Veränderung des
Stimulus, verdeutlicht werden. Verglichen mit der Bedingung Switch erreichten sie
große Effekte von d = 2,757 (Experiment I) und d = 1,891 (Experiment II). Gegenüber
der Bedingung Aufblinken ergaben sich ebenfalls große Effekte mit d = 1,926 (Experi-
ment I) und d = 1,794 (Experiment II) zugunsten der normalen Bedingung. Der Ver-
gleich der Bedingungen Switch und Aufblinken zeigte längere Dominanzzeiten bei der
Bedingung Aufblinken. Die Effektgrößen von d = -0,705 (Experiment I) und d = -0,785
(Experiment II) wiesen hier auf mittlere Effekte hin. Der Anteil an den absoluten
Inversionseffekt und Augenwechsel reduzieren Prädominanz unter binokularer Rivalität
20
Dominanzzeiten liegt in Experiment I bei 70% für die normale Bedingung, bei 14% für
Switch und bei 16% für Aufblinken. Ein fast identisches Bild zeigt sich in Experiment II
mit einer Verteilung von 69% für Normal, 15% für Switch und 16% für Aufblinken. Die
beiden experimentellen Bedingungen miteinander verglichen ergibt sich eine Verteilung
von 54:46 in Experiment I und 52:48 in Experiment II zugunsten der Bedingung Auf-
blinken.
Tabelle 5: Mittlere Dominanzzeiten (s) und Effektstärken der Switchbedingungen Experiment I
Switchbedingung MW SE d σd Cohen's d
Normal 3,481 0,275 1,881 0,682 2,757
Switch 1,600 0,227
Normal 3,481 0,275 1,603 0,832 1,926
Aufblinken 1,878 0,332
Switch 1,600 0,227 -0,278 0,394 -0,705
Aufblinken 1,878 0,332
Experiment II
Switchbedingung MW SE d σd Cohen's d
Normal 3,182 0,253 1,557 0,823 1,891
Switch 1,626 0,295
Normal 3,182 0,253 1,371 0,764 1,794
Aufblinken 1,812 0,313
Switch 1,626 0,295 -0,186 0,237 -0,785
Aufblinken 1,812 0,313
MW = Mittelwert; SE = Standardfehler; d = Mittelwert der Differenzen; σd = Standardab-weichung der Diff.; Cohen’s d= Effektgröße

Inversionseffekt und Augenwechsel reduzieren Prädominanz unter binokularer Rivalität
21
4 Diskussion
Die in Experiment I erzielten Ergebnisse lassen große Inversionseffekte sowohl für
Gesichter (d = 1,050) als auch für Häuser (d = 0,925) unter binokularer Rivalität er-
kennen (siehe Tab. 2, Seite 16). Der Inversionseffekt bei Gesichtern deckt sich mit den
bisherigen Forschungsbefunden (Bannerman et al., 2008; Engel, 1956). Dieses Ergebnis
stützt die aufgestellte Hypothese, dass aufrechte Gesichter höhere Dominanzzeiten im
Vergleich mit invertierten Gesichtern erzielen. Dieser große Effekt bleibt auch über die
weiteren Switchbedingungen Switch (d = 1,118) und Aufblinken (d = 0,965) konstant.
Der gefundene Inversionseffekt von Häusern deckt sich ebenfalls mit Befunden, dass die
aufrechte Orientierung von bedeutungsvollen Stimuli, zu denen Häuser zu zählen sind,
höhere Dominanzzeiten gegenüber ihren Inversen bewirkt (Yu & Blake, 1992).
Die Annahme, dass der Inversionseffekt von Gesichtern größer sei als der Inversions-
effekt bei Häusern, findet ebenso Bestätigung in den Ergebnissen (p < 0,05). Die ähnlich
hohe Ausprägung des Effektes überrascht allerdings und weicht von den bisherigen
Forschungsergebnissen ab (Bannerman et al., 2008).
Ein möglicher störender Einfluss könnte hierbei in der sogenannten Pareidolie liegen.
Bei diesem Phänomen handelt es sich unter anderem um die Wahrnehmung von Ge-
sichtern in Objekten oder Szenarien, in denen kein reales Gesicht existiert. Bekannte
Beispiele dafür sind der Eindruck eines Gesichts beim Anblick des Vollmondes ("Mann
im Mond") oder das vermeintliche Erkennen von Gesichtern in der Frontansicht von
Autos. Beim Betrachten einiger in dieser Studie verwendeter Hausstimuli (wie beispiels-
weise in Abb. 1 A, Seite 11) lässt sich diese Erscheinung nicht ausschließen.
Aktuelle Studien berichten von einer ähnlich starken und schnellen Aktivierung des
fusiformen Gesichtsareals während erlebter Pareidolie, wie dies sonst nur bei der
Verarbeitung von realen Gesichtern zu beobachten ist (Hadjikhani, Kveraga, Naik &
Ahlfors, 2009; Liu et al., 2014).
Sollte der Hausstimulus als gesichtsähnlich wahrgenommen werden, ist zudem nicht
auszuschließen, dass dieser als emotionaler empfunden wird als der Gesichtsstimulus.
Der neutrale Gesichtsausdruck der in dieser Studie verwendeten Gesichter wurde
allerdings bewusst gewählt, um die angenommenen Dominanzvorteile emotionaler
Gesichter zu vermeiden (Bannerman et al., 2008).
Gegen diese Theorien könnten allerdings die deutlich reduzierten Effektgrößen des
Inversionseffektes bei Häusern in den beiden experimentellen Switchbedingungen

Inversionseffekt und Augenwechsel reduzieren Prädominanz unter binokularer Rivalität
22
Switch (d = 0,413) und Aufblinken (d = 0,470) sprechen. Dieser Rückgang wäre bei
ähnlich schneller Verarbeitung und der Interpretation von Emotionen nicht zu erwarten.
Auch die großen Prädominanzvorteile von aufrechten Gesichtern im direkten Vergleich
mit aufrechten Häusern in Experiment II (d = 1,130, siehe Abb. 2 Experiment II, Seite
15) stützen diese These nicht statistisch.
Somit könnte durch diese Einflüsse der Unterschied zwischen den Inversionseffekten
von Gesichtern und Häusern unbeabsichtigt reduziert worden sein und sollte daher in
nachfolgenden Studien kontrolliert werden.
Ferner zeigte sich in Experiment I über alle Switchbedingungen hinweg eine größere
Streuung der Daten des Inversionseffektes von Gesichtern im Vergleich zu den Daten
des Inversionseffektes bei Häusern (siehe Tab. 2, Seite 16). Dies könnte unter anderem
für generell größere interindividuelle Unterschiede in der Population sprechen.
Beim direkten Vergleich von Gesichtern und Häusern unter binokularer Rivalität konnte
der angenommene Prädominanzvorteil für Gesichter belegt werden. Dabei fanden sich
große Effekte sowohl in der aufrechten (d = 1,130) als auch in der invertierten Orien-
tierung (d = 1,292). Verglichen mit den bisherigen Forschungserkenntnissen ergibt sich
allerdings wie schon in Experiment I ein ambivalentes Bild. Einerseits decken sich die
längeren Dominanzzeiten bei Gesichtern in aufrechter Orientierung mit früheren Ergeb-
nissen, der ähnlich große Effekt bei Gesichtern in invertierter Orientierung jedoch nicht
(Bannerman et al., 2008).
Anzumerken ist an dieser Stelle noch, dass sich für diese Effekte lediglich in der nor-
malen Switchbedingung statistische Bestätigung findet, denn nur dort erzielten Ge-
sichter signifikant längere Dominanzzeiten im Vergleich zu Häusern, unabhängig von
ihrer Orientierung.
Bezüglich der neuronalen Verarbeitung sprechen die bisherigen Ergebnisse für eine
Rivalität der Stimuli. Da aufrechten gegenüber invertierten Stimuli wie auch Gesichtern
gegenüber Häusern eine höhere Bedeutung der visuellen Aufmerksamkeit geschenkt
wird, lassen sich deren Prädominanzvorteile als Indiz für eine späte, sprich stimulus-
bezogene Verarbeitung betrachten.
Beide Experimente behandelten neben den stimulusspezifischen Effekten auch die
Fragestellung, ob der Wechsel eines Stimulus auf das kontralaterale Auge seine
Dominanzzeit verkürzt. Die signifikanten Vergleiche der experimentellen Switchbedin-
gungen Switch und Aufblinken erzielten sowohl in Experiment I (d = -0,705) als auch in

Inversionseffekt und Augenwechsel reduzieren Prädominanz unter binokularer Rivalität
23
Experiment II (d = -0,785) jeweils mittlere Effekte zugunsten des Aufblinkens. Der
Augenwechsel führt diesen Ergebnissen zufolge also zu einer Reduktion der Dominanz-
zeit. Dies deckt mit den Ergebnissen von Blake et al. (1980), die in ihrem Experiment
ebenfalls eine Verringerung der Dominanzzeit zeigen konnten. Allerdings ist hier der
Unterschied eher gering, verdeutlicht an der Verteilung der absoluten Dominanzzeiten
von 54:46 bzw. 52:48. Daher kann trotz signifikantem Ergebnis nicht eindeutig auf eine
Rivalität der Augen geschlossen werden.
Im Gegensatz zu Blake et al. (1980) zeigte sich hier eine bedeutende Differenz zwischen
der normalen Switchbedingung und der Bedingung Aufblinken. Diese Einbußen an
Dominanzzeit sind vermutlich auf die kurze Unterbrechung der Stimuluspräsentation
zurückzuführen, die in gleichem Ausmaß für die großen Unterschiede gegenüber der
Bedingung Switch verantwortlich sein sollten.
Als Stärken dieser Studie sei die Apparatur zu nennen. Vor allem die besonders genaue
Zeitmessung des Eingabegerätes ermöglichte eine exakte Messung der Dominanzzeiten.
Zudem lässt sich eine homogene Stichprobe bezüglich Alter und Geschlecht vorweisen.
Bei der kritischen Betrachtung dieser Studie sei zu nennen, dass in Einzelfällen die
angedachte Maximalzeit von 60 Minuten pro Sitzung überschritten wurde. Zudem zeigte
sich eine relativ große Varianz bei der Anzahl der Trials. Diese Gegebenheiten sind auf
apparative Störungen zurückzuführen und als Schwäche der Studie zu bewerten.
Aufgrund der mit zwölf Versuchspersonen nicht optimalen Stichprobengröße ist die
Interpretation der vorliegenden Ergebnisse zudem mit Einschränkung zu betrachten, da
besonders bei kleinem Stichprobenumfang die Anfälligkeit für Ausreißer erhöht ist.
Infolgedessen kann sich die tatsächliche Größe des Unterschiedes verringern bzw. ein
Effekt gänzlich verschleiert werden.
Die Ergebnisse dieser Studie konnten die besondere Rolle der Gesichtsverarbeitung an-
hand der Prädominanzvorteile von aufrechten gegenüber invertierten Gesichtern und
von Gesichtern gegenüber Häusern bestätigen. Der große Inversionseffekt bei Häusern
sollte in zukünftigen Studien auf das Phänomen der Pareidolie hin kontrolliert werden.
Für die weitere Forschung der binokularen Rivalität, die sich insbesondere mit den
Auswirkungen eines Augenwechsels auseinandersetzt, empfehlen sich "starke" Stimuli
wie emotionale, vertraute oder attraktive Gesichter, da hierbei die Auswirkungen
einzelner Bedingungen schneller sichtbar werden sollten.
Inversionseffekt und Augenwechsel reduzieren Prädominanz unter binokularer Rivalität
24
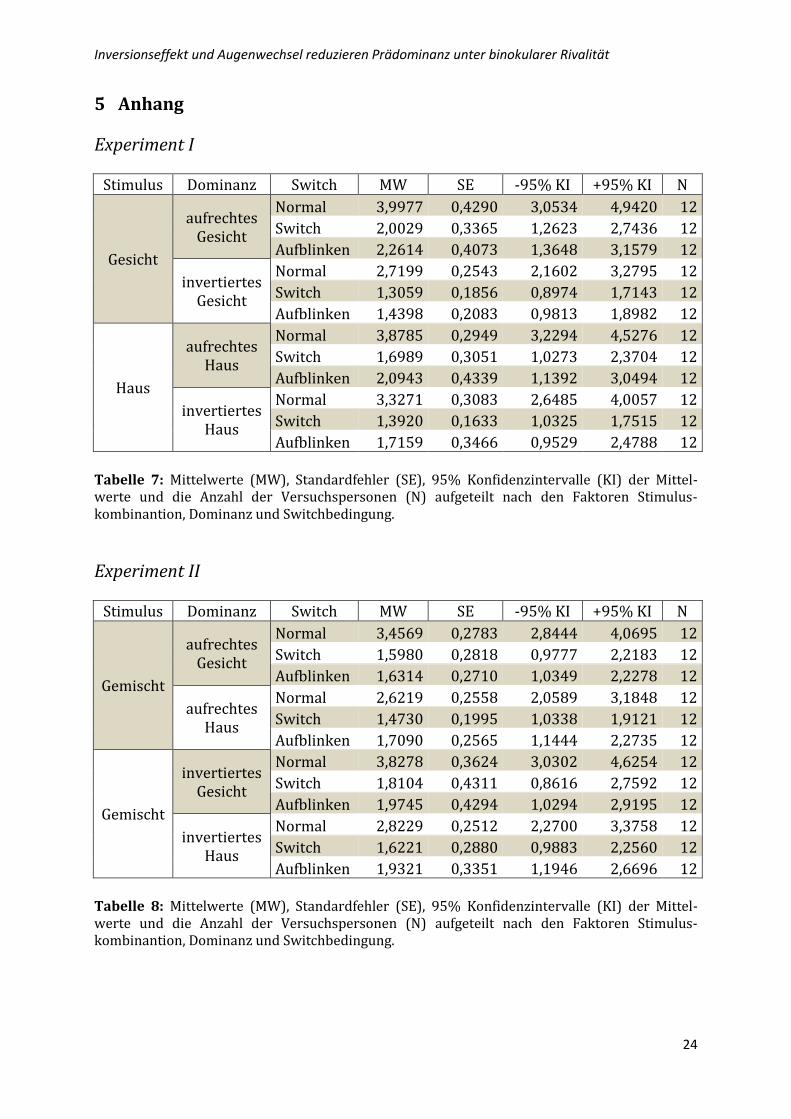
5 Anhang
Experiment I
Stimulus Dominanz Switch MW SE -95% KI +95% KI N
Gesicht
aufrechtes Gesicht
Normal 3,9977 0,4290 3,0534 4,9420 12
Switch 2,0029 0,3365 1,2623 2,7436 12
Aufblinken 2,2614 0,4073 1,3648 3,1579 12
invertiertes Gesicht
Normal 2,7199 0,2543 2,1602 3,2795 12
Switch 1,3059 0,1856 0,8974 1,7143 12
Aufblinken 1,4398 0,2083 0,9813 1,8982 12
Haus
aufrechtes Haus
Normal 3,8785 0,2949 3,2294 4,5276 12
Switch 1,6989 0,3051 1,0273 2,3704 12
Aufblinken 2,0943 0,4339 1,1392 3,0494 12
invertiertes Haus
Normal 3,3271 0,3083 2,6485 4,0057 12
Switch 1,3920 0,1633 1,0325 1,7515 12
Aufblinken 1,7159 0,3466 0,9529 2,4788 12 Tabelle 7: Mittelwerte (MW), Standardfehler (SE), 95% Konfidenzintervalle (KI) der Mittel-werte und die Anzahl der Versuchspersonen (N) aufgeteilt nach den Faktoren Stimulus-kombinantion, Dominanz und Switchbedingung.
Experiment II
Stimulus Dominanz Switch MW SE -95% KI +95% KI N
Gemischt
aufrechtes Gesicht
Normal 3,4569 0,2783 2,8444 4,0695 12
Switch 1,5980 0,2818 0,9777 2,2183 12
Aufblinken 1,6314 0,2710 1,0349 2,2278 12
aufrechtes Haus
Normal 2,6219 0,2558 2,0589 3,1848 12
Switch 1,4730 0,1995 1,0338 1,9121 12
Aufblinken 1,7090 0,2565 1,1444 2,2735 12
Gemischt
invertiertes Gesicht
Normal 3,8278 0,3624 3,0302 4,6254 12
Switch 1,8104 0,4311 0,8616 2,7592 12
Aufblinken 1,9745 0,4294 1,0294 2,9195 12
invertiertes Haus
Normal 2,8229 0,2512 2,2700 3,3758 12
Switch 1,6221 0,2880 0,9883 2,2560 12
Aufblinken 1,9321 0,3351 1,1946 2,6696 12 Tabelle 8: Mittelwerte (MW), Standardfehler (SE), 95% Konfidenzintervalle (KI) der Mittel-werte und die Anzahl der Versuchspersonen (N) aufgeteilt nach den Faktoren Stimulus-kombinantion, Dominanz und Switchbedingung.

Inversionseffekt und Augenwechsel reduzieren Prädominanz unter binokularer Rivalität
25
6 Literaturverzeichnis Bagby, J. W. (1957). A cross-cultural study of perceptual predominance in binocular rivalry. The
Journal of Abnormal and Social Psychology, 54(3), 331–334. doi:10.1037/h0046310
Bannerman, R. L., Milders, M., Gelder, B. de, & Sahraie, A. (2008). Influence of emotional facial expressions on binocular rivalry. Ophthalmic and Physiological Optics, 28(4), 317–326. doi:10.1111/j.1475-1313.2008.00568.x
Birbaumer, N., & Schmidt, R. F. (2010). Biologische Psychologie. Berlin, Heidelberg: Springer Berlin Heidelberg.
Blake, R. (1989). A neural theory of binocular rivalry. Psychological Review, 96(1), 145–167. doi:10.1037/0033-295X.96.1.145
Blake, R., & Logothetis, N. K. (2002). Visual competition. Nature Reviews Neuroscience, 3(1), 13–21. doi:10.1038/nrn701
Blake, R., Westendorf, D. H., & Overton, R. (1980). What is suppressed during binocular rivalry? Perception, 9(2), 223–231. doi:10.1068/p090223
Brascamp, J. W., van Ee, R., Pestman, W. R., & van den Berg, A. V. (2005). Distributions of alternation rates in various forms of bistable perception. Journal of Vision, 5(4), 1. doi:10.1167/5.4.1
Carter, O., Konkle, T., Wang, Q., Hayward, V., & Moore, C. (2008). Tactile rivalry demonstrated with an ambiguous apparent-motion quartet. Current biology : CB, 18(14), 1050–1054. doi:10.1016/j.cub.2008.06.027
Carter, O. L., Presti, D. E., Callistemon, C., Ungerer, Y., Liu, G. B., & Pettigrew, J. D. (2005). Meditation alters perceptual rivalry in Tibetan Buddhist monks. Current Biology, 15(11), R412. doi:10.1016/j.cub.2005.05.043
Cohen, J. (1988). Statistical power analysis for the behavioral sciences (2. ed). New York, NY: Lawrence Erlbaum Publishers.
Deutsch, D. (1974). An auditory illusion. The Journal of the Acoustical Society of America, 55(S1), S18. doi:10.1121/1.1919587
Diamond, R., & Carey, S. (1986). Why faces are and are not special: an effect of expertise. Journal of experimental psychology. General, 115(2), 107–117.
Engel, E. (1956). The rôle content in binocular resolution. The American journal of psychology, 69(1), 87–91.
Epstein, R., & Kanwisher, N. (1998). A cortical representation of the local visual environment. Nature, 392(6676), 598–601. doi:10.1038/33402
Fox, R., & Herrmann, J. (1967). Stochastic properties of binocular rivalry alternations. Perception & Psychophysics, 2(9), 432–436. doi:10.3758/BF03208783
Goldenberg, G. (2012). Visuelle Objektagnosie und Prosopagnosie. In H.-O. Karnath (Ed.), Springer-Lehrbuch. Kognitive Neurowissenschaften. Mit 28 Tabellen (3rd ed., pp. 161–171). Berlin: Springer.
Hadjikhani, N., Kveraga, K., Naik, P., & Ahlfors, S. P. (2009). Early (M170) activation of face-specific cortex by face-like objects. Neuroreport, 20(4), 403–407. doi:10.1097/WNR.0b013e328325a8e1
Kanwisher, N., & Yovel, G. (2006). The fusiform face area: a cortical region specialized for the perception of faces. Philosophical transactions of the Royal Society of London. Series B, Biological sciences, 361(1476), 2109–2128. doi:10.1098/rstb.2006.1934
Kaplan, I. T., & Metlay, W. (1964). Light intensity and binocular rivalry. Journal of Experimental Psychology, 67(1), 22–26. doi:10.1037/h0041379
Kovacs, I., Papathomas, T. V., Yang, M., & Feher, A. (1996). When the brain changes its mind: Interocular grouping during binocular rivalry. Proceedings of the National Academy of Sciences, 93(26), 15508–15511. doi:10.1073/pnas.93.26.15508

Inversionseffekt und Augenwechsel reduzieren Prädominanz unter binokularer Rivalität
26
Langner, O., Dotsch, R., Bijlstra, G., Wigboldus, Daniel H. J., Hawk, S. T., & van Knippenberg, A. (2010). Presentation and validation of the Radboud Faces Database. Cognition & Emotion, 24(8), 1377–1388. doi:10.1080/02699930903485076
Leopold, D. A., & Logothetis, N. K. (1996). Activity changes in early visual cortex reflect monkeys' percepts during binocular rivalry. Nature, 379(6565), 549–553. doi:10.1038/379549a0
Levelt, W. (1968). On binocular rivalry (Reprint). Psychological studies: Vol. 2. The Hague: Mouton.
Liu, J., Li, J., Feng, L., Li, L., Tian, J., & Lee, K. (2014). Seeing Jesus in toast: neural and behavioral correlates of face pareidolia. Cortex; a journal devoted to the study of the nervous system and behavior, 53, 60–77. doi:10.1016/j.cortex.2014.01.013
Maurer, D., Le Grand, R., & Mondloch, C. J. (2002). The many faces of configural processing. Trends in cognitive sciences, 6(6), 255–260.
McKone, E., Kanwisher, N., & Duchaine, B. C. (2007). Can generic expertise explain special processing for faces? Trends in cognitive sciences, 11(1), 8–15. doi:10.1016/j.tics.2006.11.002
Mueller, T. J., & Blake, R. (1989). A fresh look at the temporal dynamics of binocular rivalry. Biological Cybernetics, 61(3), 223-232. doi:10.1007/BF00198769
OShea, R. P., Sims, A. J., & Govan, D. G. (1997). The effect of spatial frequency and field size on the spread of exclusive visibility in binocular rivalry. Vision Research, 37(2), 175–183.
Pearson, J., Clifford, C. W., & Tong, F. (2008). The functional impact of mental imagery on conscious perception. Current Biology, 18(13), 982–986. doi:10.1016/j.cub.2008.05.048
Sheinberg, D. L., & Logothetis, N. K. (1997). The role of temporal cortical areas in perceptual organization. Proceedings of the National Academy of Sciences of the United States of America, 94(7), 3408–3413.
Tong, F., & Engel, S. A. (2001). Interocular rivalry revealed in the human cortical blind-spot representation. Nature, 411(6834), 195–199. doi:10.1038/35075583
Tong, F., Meng, M., & Blake, R. (2006). Neural bases of binocular rivalry. Trends in Cognitive Sciences, 10(11), 502–511. doi:10.1016/j.tics.2006.09.003
Tong, F., Nakayama, K., Vaughan, J., & Kanwisher, N. (1998). Binocular rivalry and visual awareness in human extrastriate cortex. Neuron, 21(4), 753–759. doi:10.1016/S0896-6273(00)80592-9
Wheatstone, C. (1838). Contributions to the physiology of vision. Part the first. On some remarkable, and hitherto unobserved, phenomena of binocular vision. Philosophical Transactions of the Royal Society of London, 128(0), 371–394. doi:10.1098/rstl.1838.0019
Wilson, H. R., Blake, R., & Lee, S. H. (2001). Dynamics of travelling waves in visual perception. Nature, 412(6850), 907–910. doi:10.1038/35091066
Yin, R. K. (1969). Looking at upside-down faces. Journal of Experimental Psychology, 81(1), 141-&.
Yu, K., & Blake, R. (1992). Do recognizable figures enjoy an advantage in binocular rivalry? Journal of Experimental Psychology: Human Perception and Performance, 18(4), 1158–1173. doi:10.1037/0096-1523.18.4.1158
Zhou, W., & Chen, D. (2009). Binaral rivalry between the nostrils and in the cortex. Current biology : CB, 19(18), 1561–1565. doi:10.1016/j.cub.2009.07.052
Software:
Inquisit: Version 4. Millisecond Software. (Experimentalsoftware)

Inversionseffekt und Augenwechsel reduzieren Prädominanz unter binokularer Rivalität
27
Erklärung für schriftliche Prüfungsleistungen
gemäß § 13, Abs. 2 und 3 der Ordnung des Fachbereichs 02 Sozialwissenschaften, Medien und Sport der Johannes Gutenberg-Universität Mainz für die Prüfung im Bachelorstudiengang B.Sc.
Psychologie vom 11. Febr. 2011, StAnz. S. 460 Hiermit erkläre ich, Brehmen, Martin 2678456 ________________________________________ Name, Vorname Matrikelnummer dass ich die vorliegende Arbeit selbstständig verfasst und keine anderen als die angegebenen Quellen oder Hilfsmittel (einschließlich elektronischer Medien und online-Quellen) benutzt habe. Mir ist bewusst, dass ein Täuschungsversuch oder ein Ordnungsverstoß vorliegt, wenn sich diese Erklärung als unwahr erweist. ________________________________________ Ort/Datum Unterschrift § 19 Abs. 3 habe ich zur Kenntnis genommen („Versucht die Kandidatin oder der Kandidat das Ergebnis einer Prüfung durch Täuschung oder Benutzung nicht zugelassener Hilfsmittel zu beeinflussen, oder erweist sich eine Erklärung gemäß § 13 Absatz 2 Satz 5 als unwahr, gilt die betreffende Prüfungsleistung als mit „nicht ausreichend“ (5,0) absolviert.“)





























