Embed Size (px)
Citation preview

HAL Id: tel-00163696https://tel.archives-ouvertes.fr/tel-00163696
Submitted on 18 Jul 2007
HAL is a multi-disciplinary open accessarchive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come fromteaching and research institutions in France orabroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, estdestinée au dépôt et à la diffusion de documentsscientifiques de niveau recherche, publiés ou non,émanant des établissements d’enseignement et derecherche français ou étrangers, des laboratoirespublics ou privés.
Transport de l’auxine et développement du noduleactinorhizien chez l’arbre tropical Casuarina glauca
Benjamin Péret
To cite this version:Benjamin Péret. Transport de l’auxine et développement du nodule actinorhizien chez l’arbre tropicalCasuarina glauca. Biologie végétale. Université Montpellier II - Sciences et Techniques du Languedoc,2007. Français. �tel-00163696�

UNIVERSITE MONTPELLIER II - SCIENCES ET TECHNIQUES DU LANGUEDOCUFR DES SCIENCES
T H E S Epour obtenir le grade de
DOCTEUR DE L'UNIVERSITE MONTPELLIER II
Discipline : Physiologie végétaleEcole Doctorale : Systèmes Intégrés en Biologie, Agronomie, Géosciences, Hydrosciences
et Environnement
présentée et soutenue publiquement par
Benjamin PERET
Le 29 juin 2007
Transport de l’auxine et développement du noduleactinorhizien chez l’arbre tropical Casuarina glauca
Directeur de thèse : Laurent LAPLAZE
JURYBruno TOURAINE PrésidentMartin CRESPI RapporteurPhilippe NORMAND RapporteurDavid BARKER ExaminateurLaurent NOEL ExaminateurLaurent LAPLAZE Examinateur

“Patience et longueur de tempsFont plus que force ni que rage.”
Jean de la Fontaine ( Les Fables , Livre II, Fable 11)

REMERCIEMENTSJe tiens à exprimer ma plus profonde gratitude, ma plus chaleureuse amitié ainsi que mon plus
grand respect au Dr. Laurent LAPLAZE qui a assuré la direction de ma thèse. Ces trois années passées àses côtés ont été riches en enseignement, en travail efficace et également en franche rigolade. Merci dem’avoir permis de commencer ma carrière de chercheur dans de si bonnes conditions.
Je tiens à assurer le renouvellement de mon plus grand respect à l’égard du Dr. Didier BOGUSZ.J’adresse à l’ensemble des membres, passés et présents, de l’équipe Rhizogénèse mes
remerciements les plus sincères pour l’aide qu’ils ont pu m’apporter et la convivialité dont ils ont faitpreuve. Merci donc au Dr. Claudine FRANCHE pour ses conseils en transgénèse végétale, au Dr. ValérieHOCHER pour son aide en QRT-PCR, à l’ingénieur d’études Florence AUGUY pour son aide en biologiemoléculaire, au Dr. Sergio SVISTOONOFF pour ses conseils sur le présent manuscrit, au Dr. PatrickDOUMAS pour son soutien pendant la rédaction et ses conseils sur le manuscrit, au Dr. FabienneCARTIEAUX, au Dr. Hassen GHERBI, au Pr. Christian JAY-ALLEMAND et au doctorant Mikaël LUCAS.
Je remercie tout particulièrement le Pr. Malcolm BENNETT de l’université de Nottingham de m’avoiraccueilli dans son laboratoire. Je le remercie également pour ses nombreux conseils scientifiques et sasympathie légendaire. Mes remerciements s’adressent également aux membres de son équipe et entout premier lieu au Dr. Ranjan SWARUP.
Je salue mes deux stagiaires Leen JANTSEN et Gaëlle DEVOS qui ont toutes deux participé à ce projetde thèse avec beaucoup de rigueur, d’efficacité et de sympathie.
Je remercie Myriam COLLIN qui m’a apporté une aide des plus précieuses en histologie et qui aformé mes deux stagiaires au maniement du microtome et autres techniques d’histologie.
Je remercie Martine BANGRATZ pour son aide et son soutien au cours de mes longs repiquages decals. Je remercie l’ensemble des membres de GeneTrop pour leur aide ponctuelle ou récurrente.
J’adresse au Dr. Laurent NUSSAUME toutes mes excuses et promets de lui offrir une gaufre belge(avec supplément) dès que l’occasion s’en présentera.
Je salue toute l’équipe d’enseignement du module “Bases de la physiologie végétale” de L2 danslequel j’ai effectué mon monitorat. Merci au Dr. Michel COUPE, au Dr. Pierre CZERNIC, au Dr. LaurenceMARQUES, au Dr. Claire CORRATGE, à la doctorante Cécile RANGIN, au doctorant Karl RAVET et à la trèsefficace Lydia GAMET. J’adresse mes salutations à tous les moniteurs que j’ai rencontrés au cours despassionnantes formations dispensées par le Centre d’Initiation à l’Enseignement Supérieur. Je penseévidemment aux quelques 600 étudiants à qui j’ai tenté d’enseigner les bases de la physiologievégétale en espérant que certains auront suivi le bon chemin.
Je remercie le Pr. Philippe NORMAND, le Pr. Bruno TOURAINE, le Dr. David BARKER, le Dr. MartinCrespi et le Dr. Laurent NOEL d’avoir accepté de faire partie de mon jury de thèse.
Je présente par avance mes excuses aux personnes que j’aurais oublié de remercier.
Je dédie ce travail de thèse à mon épouse Caroline et à l’enfant qu’elle porte.

TABLE DES MATIÈRES
ABRÉVIATIONS 1
TABLE DES FIGURES 2
ETUDE BIBLIOGRAPHIQUE 3
I. Introduction générale 3
II. Symbioses Rhizobium-Légumineuses 41) Généralités 42) Le partenaire bactérien 53) La formation du nodule 5
III. Les symbioses actinorhiziennes 91) Généralités 92) Aspects phylogénétiques et évolutifs 93) Le partenaire bactérien 94) La formation du nodule actinorhizien 10
IV. Formation de la racine latérale 141) Généralités 142) Initiation 153) Organisation du primordium 174) Émergence 17
V. L’hormone végétale auxine 181) Introduction 182) Perception et signalisation 183) Homéostasie 184) Transport 20
VI. Objectifs de la thèse 211) Isolement et caractérisation fonctionnelle des gènes AUX-LAX chez Casuarina glauca 222) Implication au cours du développement du nodule actinorhizien 223) Rôle des transporteurs d’influx dans le développement : exemple de LAX3 22
PARTIE I - ISOLEMENT ET CARACTÉRISATION DE GÈNES AUX-LAX CHEZ CASUARINAGLAUCA 23
I. Isolement d’homologues de gènes AUX-LAX chez C. glauca 231) Obtention des ADNc et des séquences génomiques 232) CgAUX1 et CgLAX3 appartiennent à une petite famille de gènes 243) Structure des gènes CgAUX1 et CgLAX3 244) Séquences protéiques déduites 25
II. Comparaison de l’expression des gènes AtAUX-LAX et CgAUX-LAX 26
III. Analyse fonctionnelle 271) Complémentation fonctionnelle du mutant aux1 282) Complémentation fonctionnelle du mutant lax3 29
IV. Discussion 311) Une petite famille multigénique AUX-LAX chez C. glauca 31

2) Divergence fonctionnelle dans la famille AUX-LAX 313) Bases moléculaires de la divergence 32
PARTIE II - ÉTUDE DU RÔLE DU TRANSPORT D’INFLUX AU COURS DUDÉVELOPPEMENT DU NODULE ACTINORHIZIEN 33
I. Introduction 33
II. Analyse de l’expression de CgAUX1 et CgLAX3 331) Expression non-symbiotique 332) Expression au cours de la symbiose 343) CgAUX1 et CgLAX3 ne sont pas inductibles par l’auxine 35
III. L’inhibition du transport d’influx d’auxine perturbe le processus de nodulation 36
IV. Dosages d’auxine 381) Production d’auxine par la bactérie Frankia 382) Accumulation d’auxine au cours de la nodulation 39
V. Discussion 391) L’influx d’auxine est associé au mécanisme d’infection par Frankia 392) CgAUX1 pourrait créer un puits d’auxine au site d’infection 403) CgAUX1 n’est pas impliqué dans l’organisation du primordium nodulaire 40
PARTIE III - CIBLES DE LA SIGNALISATION AUXINIQUE DEPENDANTE DU TRANSPORTD’INFLUX 42
I. Introduction 421) AtLAX3 est impliqué dans l’émergence de la racine latérale 422) AtLAX3 est exprimé dans les cellules en face du primordium 423) AtLAX3 est inductible par l’auxine 434) Conclusion 44
II. AtLAX3 permet le remodelage de la paroi dans les cellules de l’épiderme et du cortexau cours de l’émergence 44
1) L’induction d’AIR3 par l’auxine dépend de LAX3 442) Un ensemble de gènes de remodelage de la paroi induits par l’auxine et dépendants de
LAX3 453) Modèle d’action d’AtLAX3 dans l’émergence de la racine latérale 46
III. Influx d’auxine et remodelage de la paroi au cours de l’infection par Frankia 471) Régulation de Cg12 par un signal auxinique 472) Autres gènes de remodelage de la paroi chez Casuarina glauca 48
IV. Discussion 491) Amplification du signal auxinique par l’influx d’auxine 492) Nature du signal symbiotique 50
CONCLUSION GÉNÉRALE – PERSPECTIVES 51
I. Le signal auxinique permet l’infection de la cellule végétale par des micro-organismes 51
II. Le nodule actinorhizien : une racine latérale modifiée ? 53
MATÉRIEL ET MÉTHODES 55

I. Matériel 551) Matériel végétal 552) Matériel bactérien 553) Matériel nucléique 55
II. Méthodes 551) Milieux de culture (plantes) 552) Milieux de culture (bactéries) 573) Transformation génétique des plantes 584) Transformation génétique des bactéries 595) Biologie moléculaire 606) Histologie 617) Analyses phénotypiques 628) Dosages 62
RÉFÉRENCES BIBLIOGRAPHIQUES 63
RÉSUMÉ 71
ABSTRACT 71

1
ABRÉVIATIONS
α32P-dCTP : désoxy cytidinetriphosphate marquée au 32P en position α
2,4D : acide 2,4 dichlorophénoxyacétique
35S : promoteur 35S du virus de lamosaïque du chou-fleur
ABC : ATP binding cassetteADN : acide désoxyribonucléiqueADNc : ADN complémentaireADP : adénosine diphosphateARF : auxin response factorARF/GEF : ADP ribosylation
factor/GTPase exchange factorARN : acide ribonucléiqueARP : auxin repressed proteinATP : adénosine triphosphateAux/IAA : auxin/indole-3-acetic proteinAuxRE : auxin response elementBAP (milieu) : Buffered mineral medium
Added of PhosphatidylcholineBD (milieu) : milieu de Broughton et
DilworthCDK : cyclin dependent kinaseEDTA : acide éthylène
diaminotétraacétiqueEST : expressed sequence tagGFP : green fluorescent proteinGUS : β-glucuronidase
HPLC : high performance liquidcrhomatography
HRGP : hydroxyproline rich glycoproteinIAA : acide indole acétiqueiPA : isopentényl adénosineLB (milieu) : milieu liquide de Luria et
BertaniMATAB : mixed alkyl trimethyl
ammonium bromideMDR/PGP : mulidrug resistance/P
glycoproteinNAA : acide naphthalène acétiqueNOA : acide naphtoxy acétiqueORF : open reading framePAA : acide phénylacétiqueRACE PCR : rapid amplification of cDNA
ends by PCRRT-PCR : reverse transcription-
polymerase chain reactionSDS : dodécylsulfate de sodiumSSC : citrate de sodium, chlorure de
sodiumTris : trishydroxyméthylaminométhaneUTR : untranscribed regionX-Glc : 5-bromo-4-chloro-3-indolyl-
D-glucuronideYFP : yellow fluorescent protein

2
TABLE DES FIGURESFigure 1. Cycle de l’azoteFigure 2. Structure de la nitrogénaseFigure 3. Phylogénie des LégumineusesFigure 4. Structure des facteurs NodFigure 5. Déformation des poils racinaires chez le haricotFigure 6. Modèle de la voie de transduction du signal symbiotique chez le lotierFigure 7. Infection intracellulaire chez les LégumineusesFigure 8. Types nodulaires chez les LégumineusesFigure 9. Infection intracellulaire et différenciation du symbiosome chez les LégumineusesFigure 10. Filaments de FrankiaFigure 11. Infections intra et extra-cellulaire chez les plantes actinorhiziennesFigure 12. Processus d’infection et organogenèse d’un lobe nodulaire chez les plantes
actinorhiziennesFigure 13. Nodule et racines nodulaires de Casuarina glaucaFigure 14. Différents stades de formation du primordium de racine latérale chez Arabidopsis
thalianaFigure 15. Anatomie de la racine d’Arabidopsis thalianaFigure 16. Régulation du cycle cellulaire dans le péricycleFigure 17. Implication des flux d’auxine dans les processus développementauxFigure 18. Formules chimiques des différents types d’auxineFigure 19. Signalisation auxiniqueFigure 20. Homéostasie de l’auxineFigure 21. Voies de biosynthèse de l’auxineFigure 22. Transport de l’auxine dans l’apex racinaireFigure 23. La famille AUX-LAX de Casuarina glaucaFigure 24. Comparaison des séquences protéiques déduites de CgAUX1 et CgLAX3Figure 25. Profils d’expression d’AtAUX1, CgAUX1, AtLAX3 et CgLAX3 chez Arabidopsis thalianaFigure 26. Relation phylogénique entre Casuarinacées et BrassicacéesFigure 27. Phénotype gravitropique des mutants aux1-22 complémentés avec les gènes de
Casuarina glaucaFigure 28. Détection des ARNm de CgLAX3 chez les mutants aux1-22 complémentés par CgLAX3Figure 29. Analyse du phénotype de sensibilité/résistance à l’auxineFigure 30. Densité de racines latérales chez le mutant lax3 complémenté par CgLAX3Figure 31. Nombre de racines latérales chez le mutant aux1lax3 complémenté par CgLAX3Figure 32. AXR4 est nécessaire pour permettre la localisation membranaire d’AUX1Figure 33. Détection des ARNm de CgAUX1 et CgLAX3Figure 34. Expression non-symbiotique de CgAUX1 et CgLAX3Figure 35. Expression de CgAUX1 au cours de l’infectionFigure 36. Expression de CgLAX3 dans le nodule actinorhizien chez Allocasuarina verticillataFigure 37. Induction de CgAUX1 et CgLAX3 par les auxinesFigure 38. Le 1-NOA retarde la nodulation en inhibant le transporteur d’influx d’auxine CgAUX1Figure 39. Dosage des auxines produites par Frankia et Casuarina glaucaFigure 40. Phénotype du mutant lax3Figure 41. Profil d’expression d’AtLAX3Figure 42. AtLAX3 est induit par l’auxine de façon dépendante de SLR1/IAA14Figure 43. PIN2 permet le transport basipète de l’auxine dans le primordiumFigure 44. AtAIR3 est induit par l’auxine de façon dépendante d’AtLAX3Figure 45. Gènes de remodelage de la paroi induits par l’auxine de façon dépendante d’AtLAX3Figure 46. Modèle du rôle d’AtLAX3 au cours de l’émergenceFigure 47. Effet du NOA sur l’expression de Cg12, CgAUX1 et CgHbFigure 48. Induction par l’auxine et expression en réponse à l’infection des gènes de remodelage
de la paroiFigure 49. Modèle du rôle de CgAUX1 au cours de l’infection par FrankiaFigure 50. Processus de mise en place des symbioses mycorhiziennesFigure 51. Construction des vecteurs binaires ProCgAUX1:GUS, ProCgLAX3:GUS et ProAtAUX1:CgAUX1Figure 52. Construction des vecteurs binaires ProAtAUX1:CgLAX3 et ProAtLAX3:CgLAX3

3
ETUDE BIBLIOGRAPHIQUE
I. Introduction généraleL’azote est le premier élément minéral
limitant la croissance des plantes car lesseules formes assimilables sont présentesen faible quantité dans les sols. Lamajeure partie de l’azote se trouve sousforme de diazote atmosphérique, maisseules quelques espèces de procaryotesont la possibilité de l’utiliser pour leurnutrition azotée. On distingue cettefixation biologique de l’azote (environ1014g d’azote par an) et la fixationindustrielle selon le procédé d’Haber-Bosch (environ la moitié de la fixationbiologique) pour la production d’engraisazotés (Arp, 2000). Dans un systèmebiologique fixateur d’azote, les conditionsoptimales de la catalyse biologiquecorrespondent à une pression de 0,2 à 1,0atm de N2 et une température de 30-35°C,alors que les conditions de la catalysechimique sont très sévères : pression de250 à 1000 atm de N2 et température de450°C (Hardy et Knight, 1968).
L’apport de nitrate (NO3-) dans les sols,
source principale d’azote pour les plantes,est réalisé par oxydation de l’ammonium(NH4
+) en nitrite (NO2-) puis en nitrate par
des bactéries chimiolithotrophes
(Nitrosomonas et Nitrobacter) selon unprocessus nommé nitrification, favorisé
par une température élevée et un sol bienaéré (processus aérobie). La dénitrificationqui appauvrit les sols en nitrate estréalisée par d’autres bactéries(Pseudomonas, Thiobacillus) en conditionsanaérobies (Figure 1).
La fixation biologique de l’azoteatmosphérique est catalysée par uncomplexe enzymatique appelé complexenitrogénase (Rees et Howard, 2000). Cetteenzyme a été mise en évidenceuniquement chez des procaryotes. Lecomplexe nitrogénase est très conservéchez les bactéries fixatrices d’azote tantau niveau de sa séquence que de sastructure. Il est constitué de deuxmétalloprotéines : le site de la réduction
du substrat est la protéine MoFe(composant I ou dinitrogénase) et ledonneur d’électrons est la protéine Fe(composant II ou dinitrogénase réductase).Le composant I est un hétérotétramère detype α2β2 codé par les gènes nifD et nifK etle composant II est un homodimère codépar le gène nifH (Figure 2 ; Howard etRees, 2000 ; Rangaraj et al, 2000).
La régulation de l’expression de lanitrogénase est complexe ; elle fait
Figure 1. Cycle de l’azoteLa grande majorité de l’azote se trouve sous forme dediazote (N2) non-assimilable par les plantes. La fixationbiologique de l’azote par les bactéries en symbiose avecles plantes actinorhiziennes et les Légumieuses permetd’accéder à une source quasi-illimitée d’azote.
Figure 2. Structure de la nitrogénaseLe complexe nitrogénase catalyse la réaction de fixation del’azote atmosphérique. Cette enzyme n’existe que chez lesorganismes procaryotes. Elle est constituée de deuxcomposants : le composant I est un hétérotétramèreconstitué de deux sous-unités α représentées en bleu et vertet deux sous-unités β représentées en rouge et orange (A), ledétail de la sous-unité α est donné (B) et le composant II estun homodimère (C). D’après Rubio et Ludden, 2005.

4
intervenir au niveau transcriptionneld’autres gènes nif de l’opéron (nifA et nifLnotamment) et au niveau post-traductionnel un système d’ADP-ribosylation (répression de l’enzyme parNH4
+ et l’obscurité). Le complexenitrogénase est extrêmement labile enprésence de dioxygène (Halbleib etLudden, 2000). La réaction catalysée parcette enzyme est la suivante :
N2 + 8H+ +8e- + 16ATP → 2NH3 + H2+ 16 ADP + 16 Pi
La réaction de fixation de l’azote esttrès coûteuse en énergie (ATP et pouvoirréducteur). De ce fait, la fixation de l’azotepar les bactéries diazotrophes à l’état libreest peu efficace : de l’ordre de la dizainede kg N.ha-1.an-1. L’associationsymbiotique entre des bactéries fixatricesd’azote et certaines plantes permetd’améliorer considérablement cettevaleur pour atteindre une centaine de kgN.ha-1.an-1 (Bohlool et al, 1992). Ondistingue plusieurs types de symbiosesfixatrices d’azote, les principaux typessont les symbioses nodulaires(actinorhiziennes et Légumineuses) et lessymbioses avec les cyanobactéries. Lessymbioses avec les cyanobactéries neconduisent pas à proprement parler à laformation de nouveaux organesspécialisés dans la symbiose mais plutôtau détournement d’organes existants :présence d’une cavité chez Azolla abritantla bactérie Anabaena, infection de glandessymbiotiques par Nostoc chez Gunnera etracines coralloïdes infectées par Nostocchez le Cycas. À l’inverse, les symbiosesnodulaires sont des associations trèsétroites puisqu’elles nécessitent laformation d’un nouvel organe végétal : lenodule, qui héberge la bactérie et au seinduquel ont lieu les échanges entre lesdeux symbiontes. La symbioseRhizobium-Légumineuses est de loin laplus étudiée car elle concerne beaucoupd’espèces d’intérêt agronomique(alimentation humaine et animale). Lessymbioses actinorhiziennes sont moinsétudiées mais ont néanmoins une grandeimportance écologique.
II. Symbioses Rhizobium-Légumineuses
1) GénéralitésLes Légumineuses représentent une
superfamille chez les angiospermes,comprenant plus de 650 genres et 18000espèces. Elles sont divisées en trois sous-familles : les Mimosoïdées, lesCaesalpinoïdées et les Papilionoïdées(Figure 3). De nombreuses Légumineusesconstituent une source majeure de
protéines et d’huiles végétales (Graham etVance, 2003) et sont largement cultivéessur l’ensemble de la planète. On peut citerpar exemple : le haricot (Phaseolusvulgaris), le soja (Glycine max), le pois(Pisum sativum), le pois chiche (Cicerarietinum), la fève (Vicia faba), le niébé(Vigna unguiculata), la lentille (Lensesculenta), la cacahuète (Arachis hypogea).La plus grande partie des Légumineuses(88% des espèces étudiées ; de Faria et al,1989) interagissent avec Rhizobium pourformer des nodules fixateurs d’azote(Hirsch et al, 2001). De ce fait, lesLégumineuses sont parmi les plantes lesplus étudiées (Patriarca et al, 2004 ; Gage,2004 ; Stacey et al, 2006). Notamment,l’émergence de deux plantes modèles : lelotier (Lotus japonicus ; Handberg etStougaard, 1992 ; Udvardi et al, 2005) etla luzerne (Medicago truncatula ; Barker etal, 1990) a permis d’accélérer l’étude desmécanismes de mise en place de lasymbiose.
Figure 3. Phylogénie des LégumineusesLes Légumineuses sont groupées en trois grandes sous-familles : les Mimosoïdées, les Caesalpinoïdées et lesPapilionoïdées, comprenant de nombreuses espècescultivées. D’après Udvardi et al. 2005.

5
2) Le partenaire bactérienLe terme « rhizobia » regroupe des
bactéries appartenant aux quatresdifférentes branches de la sous-classe αdes protéobactéries : la brancheMesorhizobium-Sinorhizobium-Rhizobium, la branche desBradyrhizobium, la branche desAzorhizobium et la branche desMethylobacterium (Moulin et al, 2001). Cesont des bactéries en forme de bâtonnetsà l’état libre, non sporulantes etgénéralement mobiles grâce à la présenced’un ou plusieurs flagelles. Le groupe desrhizobia comprend des bactéries trèsdiverses qui ont en commun leur aptitudeà induire la formation de nodules fixateursd’azote sur les racines ou les tiges desLégumineuses. Elles n’acquièrent engénéral leur capacité à fixer l’azoteatmosphérique qu’au sein des nodules (àl’exception de Bradyrhizobium etAzorhizobium).
3) La formation du noduleLes conditions requises avant la mise
en place de la symbiose sont une faibleteneur en azote du sol et unephotosynthèse active pour assurer unesource suffisante d’énergie (Kondorosi etKondorosi, 2000).
a) PréinfectionL’interaction entre la plante et la
bactérie débute dans la rhizosphère, laplante permettant la croissance desbactéries de manière sélective (Savka et al,2002). Les rhizobia sont attirés vers lespoils racinaires par une large gamme desubstances, principalement par lesphénylpropanoïdes exsudés par la racine(Kape et al, 1991). Une production plusimportante de ces composés est observéeen condition de carence azotée (Coronadoet al, 1995). Les flavonoïdes présentsdans les exsudats racinaires induisentl’expression des gènes nod bactériens quigouvernent la production des facteursNod, des lipochitooligosaccharides dontles décorations variables déterminent laspécificité d’hôte (Figure 4 ; Perret et al,2000). Les facteurs Nod induisent desévénements morphologiques,
physiologiques et moléculaires chez laplante hôte.
La déformation du poil racinaire estobservée environ 12 à 24 heures aprèsinoculation chez M. sativa (Wais et al,2002). Elle fait intervenir des changementsdans l’arrangement des microtubules(Timmers et al, 2007). Plus précisément,les poils racinaires peuvent adopterdifférentes formes en fonction de leurstade de développement (Figure 5) : encrosse de berger, courbés, renflés,entrelacés, déformés, branchés ou joints(Wood et Newcomb, 1989).
La réponse de la plante fait intervenir :i) des variations de la concentrationintracellulaire en calcium (Supanjani et al,2006), ii) la production de composés quine sont pas des flavonoïdes et qui
Figure 4. Structure des facteurs NodStructure chimique d’un facteur Nod générique avecquelques substitutions observées chez les différentesespèces de rhizobia indiquées en couleur. Les variationsles plus courantes de la chaine d’acides gras sontégalement représentées. Ac, acétyl ; Ara, arabinosyl ;Cb, carbonyl ; Fuc, fucosyl ; Me, méthyl ; S, sulfuryl.D’après Cullimore et al., 2001.
Figure 5. Déformation des poils racinaires chez le haricotDifférents types de déformation des poils racinaires sontobservés chez le haricot (Phaseolus vulgaris) après contactavec la bactérie Rhizobium etli. Gauche : différentes zonescorrespondant aux différents stades de différenciation despoils racinaires. Droite : types de déformations courammentobservées en réponse à la bactérie symbiotique. D’aprèsPatriarca et al., 2004.

6
augmentent l’induction des gènes nodbactériens (van Brussel et al, 1990), iii)l’expression du gène de la chalconesynthase, enzyme clé de la voie debiosynthèse des flavonoïdes (Yang et al,1992), iv) la production de chitinases quidégradent les facteurs Nod et contrôlentleur concentration (Ovtsyna et al, 2005), v)des changements dans la balancehormonale (Mathesius et al, 2000a) et vi)la production d’espèces réactives del’oxygène (Baier et al, 1999). Certainsgènes impliqués dans la voie detransduction du signal ont été identifiés.Parmi eux, on peut citer un canalcationique (DMI1 ; Ané et al, 2004), unekinase de type récepteur (DMI2 ; Endre etal, 2002), une kinase dépendante decalmoduline/calcium (DMI3 ; Mitra et al,2004) et des régulateurs de transcription(NSP1 et NSP2 ; Kaló et al, 2005 ; Smit etal, 2005) chez M. truncatula (Figure 6).
Enfin, le récepteur SymRK du lotier(homologue de DMI2) intervient égalementdans cette voie de transduction puisqueles mutants symrk sont bloqués au coursde la phase d’infection du poil racinaire(Capoen et al, 2005). Les mutants dmi1,dmi2 et dmi3 de M. truncatula sontincapables de former des mycorhizes ce
qui démontre l’existence de voies designalisation communes entre ces deuxtypes de symbioses.
Néanmoins, les mécanismes d’actionne sont pas encore tous identifiés.Notamment, le récepteur des facteurs Nodest toujours inconnu (Cullimore et al, 2001; Geurts et al, 2005). Deux kinases LysM àdomaine de type récepteur pourraientjouer ce rôle chez le lotier (NFR1 et NFR5 ;Madsen et al, 2003 ; Radutoiu et al, 2003)et chez M. truncatula (NFP et LYK3 ; Amoret al, 2003 ; Limpens et al, 2003). En effet,les premiers événements observés dans lepoil racinaire en réponse à l’application defacteurs Nod (influx de calcium etoscillations calciques) sont absents chezles mutants nfr1 et nfr5 (Miwa et al, 2006).
b) InfectionOn distingue deux types d’infection : la
voie intracellulaire qui est la plus étudiéeet la voie intercellulaire (ou « crack-entry »). Au cours de l’infectionintracellulaire, la pénétration de la bactérieest facilitée par la courbure du poilracinaire qui crée une zone confinée danslaquelle la bactérie est entourée par laparoi végétale (Figure 7). Un cordon
d’infection est initié à partir de ce pointpar hydrolyse de la paroi (Mateos et al,2001), invagination de la membranevégétale et production de matériel pariétalpar la plante (Gage et Margolin, 2000 ;Gage, 2004). Le cordon d’infection est unestructure tubulaire qui croît à l’intérieur dela cellule et dans laquelle la bactérieprolifère. L’infection d’une cellule corticale
DMI3/
Figure 6. Modèle de la voie de transduction du signalsymbiotique chez le lotierDe nombreux composants de la voie de transduction du signalont été identifiés chez les Légumineuses. Notamment, les deuxkinases à domaine récepteur NFR1 et NFR5 sont impliquéesdans la reconnaissance spécifique des facteurs Nod. Lesmessagers secondaires incluent des phospholipases C et D (PLCet PLD) et des changements de la concentration cytosolique etnucléaire en calcium. L’activation d’une kinase via unecalmoduline entraine l’expression des gènes de nodulines parl’intermédiaire des régulateurs transcriptionnels NSP1 et NSP2.D’après Oldroyd et Downie, 2006.
Figure 7. Infection intracellulaire chez les LégumineusesLa bactérie colonise la rhizosphère et entre en contact avec lepoil racinaire (A). Les facteurs Nod induisent la courbure dupoil racinaire ce qui permet la création d’une zone confinéepour la bactérie qui initie l’infection (B). La mise en place ducordon d’infection suit le déplacement du noyau vers la basedu poil (C). D’après Perret et al., 2000.

7
par le cordon d’infection est précédéed’un réarrangement du cytosquelette,d’un déplacement du noyau vers le centrede la cellule et de la formation d’unestructure similaire à un phragmoplaste(van Brussel et al, 1992).
L’infection intercellulaire se faitgénéralement au niveau de passageslibérés par l’émergence des racineslatérales ou adventives, ou bien parfoisdirectement à travers la lamelle moyenneentre deux cellules du rhizoderme(Pawlowski et Bisseling, 1996). Lesrhizobia progressent ensuite vers leprimordium nodulaire de manièreintercellulaire ou deviennentintracellulaires en formant des cordonsd’infection.
c) Développement du noduleL’infection de la plante par les rhizobia
induit la dédifférenciation et la division descellules du cortex (Foucher et Kondorosi,2000). Les nodules de type indéterminé(M. truncatula, Pisum sativum) sont formésà partir du cortex interne alors que lesnodules de type déterminé (L. japonicus,Glycine max, Phaseolus vulgaris) sontformés à partir du cortex externe. Lapersistance du méristème chez lesespèces à nodules de type déterminé esttrès éphémère et la croissance enlongueur du nodule est limitée. Unecroissance en épaisseur a lieu parhypertrophie des cellules corticales et pardes divisions de cellules contenant déjàdes Rhizobia. Ce processus de formationse traduit par une forme sphérique et un
état de différenciation identique pourtoutes les cellules. Dans le cas des espècesà nodules de type indéterminé, la zoneméristématique est persistante ce qui setraduit par une forme allongée (Figure 8).
Dans les deux cas, les cellules ducortex se divisent de manière anticlinepuis péricline (Timmers et al, 1999).Toutes les cellules du cortex ne se divisentpas, ce qui semble indiquer que lasusceptibilité de ces cellules pourrait êtreliée à un statut particulier, notamment unemodification de la concentration enhormones (Mathesius et al, 2000a). Demanière concomitante, les cellulesvoisines développent des cordons depréinfection, constitués de pontscytoplasmiques alignés de façon radiale(van Brussel et al, 1992). Ces structuresguident la croissance des cordonsd’infection en direction du primordiumnodulaire en formation.
L’utilisation d’inhibiteurs du transportd’efflux d’auxine entraîne la formation de« pseudonodules » (Fang et Hirsch, 1998)suggérant un rôle de l’auxine dans laformation du nodule. De plus, les facteursNod produits par Rhizobium avantl’infection entraînent une modification dela balance hormonale de la plante. Lesmécanismes moléculaires responsables deces changements sont inconnus mais ilsemble que les facteurs Nod agissent surles flux d’auxine à deux niveaux : uneinhibition du transport de l’auxine(Mathesius et al, 1998 ; Boot et al, 1999)et l’induction de la synthèse deflavonoïdes (Mathesius et al, 2000a). Lesflavonoïdes sont susceptibles deprovoquer l’accumulation de l’auxine enréduisant son oxydation de manièredirecte (substrats de l’enzyme) ouindirecte (en réagissant avec H2O2 ;Mathesius, 2001). Leur rôle dansl’inhibition du transport de l’auxine estégalement supposé (Brown et al, 2001).Des travaux récents montrent quel’extinction par la technique d’ARNinterférent de l’expression du gène de lachalcone synthase (CHS) chez M.truncatula entraine une augmentation dutransport de l’auxine associée à uneinhibition de la nodulation (Wasson et al,2006).
pn
Figure 8. Types nodulaires chez les LégumineusesNodule de type indéterminé présentant une zoneméristématique (I), une zone de préfixation (II), une zonede fixation (III) et une zone de sénescence (IV) (A). Nodulede type déterminé (B). pn : parenchyme nodulaire. D’aprèsPawlowski et Bisseling, 1996.

8
L’utilisation de promoteurs induits parl’auxine (GH3) a permis de montrer quel’auxine s’accumule dans les premièrescellules du cortex qui vont former leprimordium nodulaire (Mathesius et al,2000b). Le transport d’auxine dans lenodule en formation implique les gènes detransport d’influx chez M. truncatula lorsde la formation du primordium nodulaireet plus tard au cours de la différenciationdu système vasculaire (de Billy et al, 2001).
Les cytokinines jouent également unrôle dans la formation des nodules.L’utilisation d’un promoteur d’Arabidopsisexprimé en réponse aux cytokinines(ARR5 ; D'Agostino et al, 2000) a permisde montrer une accumulation précoce,dans le poil racinaire infecté et plus tarddans le primordium nodulaire chez lelotier (Lohar et al, 2004). Le récepteurputatif des cytokinines a été identifié chezMedicago par homologie avec le récepteurCRE1 d’Arabidopsis (Gonzalez-Rizzo et al,2006). L’extinction de son expression parla technique d’ARN interférent entraineune inhibition de la nodulation chezMedicago (Gonzalez-Rizzo et al, 2006)alors qu’une mutation gain de fonctionprovoque la formation spontanée denodules en l’absence de bactériessymbiotiques chez le lotier (Tirichine et al,
2007).L’éthylène joue également un rôle dans
la nodulation. En effet le mutant sickle deM. truncatula qui est insensible àl’éthylène présente un phénotyped’hypernodulation (Prayitno et al, 2006).Ce phénotype est associé avec uneaugmentation du transport de l’auxine,principalement le transport d’efflux. Chezla plupart des légumineuses, l’effetinhibiteur de l’éthylène sur la nodulation aété démontré (Lee et Larue, 1992 ;Schmidt et al, 1999 ; Nukui et al, 2000).L’éthylène jouerait un rôle dans lepositionnement des nodules en face despôles de xylème (Heidstra et al, 1997).
d) Structure du noduleL’infection du nodule indéterminé se
fait par sa base, ce qui établit un gradientde différenciation et définit plusieurszones :• la zone méristématique (zone I)
située à l’apex. Cette zone est toujoursdépourvue de bactéries.
• la zone de préfixation (zone II) quicontient les cellules corticalesnouvellement produites par leméristème et qui sont envahies pardes cordons d’infection rhizobiens. Lesbactéries sont déversées dans lescellules, entourées par la membranepéribactéroïdienne, et leurdifférenciation en bactéroïdescommence (Figure 9). À ce stade, ellesne fixent pas encore l’azote.
• l’interzone II-III dans laquelle ladifférenciation des bactéroïdes sepoursuit et la fixation de l’azotecommence. Cette zone se caractérisepar la présence de nombreuxamyloplastes.
• la zone de fixation (zone III) où lesbactéroïdes pleinement différenciésfixent activement l’azote.
• la zone de sénescence (zone IV) quiest présente chez les nodules âgés.
Les nodules de Légumineusesprésentent une structure similaire à celled’une tige avec les tissus vasculairespériphériques qui se raccordent à ceux dela racine et une zone centrale infectée par
Figure 9. Infection intracellulaire et différenciation dusymbiosome chez les LégumineusesLes bactéries sont déversées dans la cellule végétale parfusion des vésicules lipidiques avec l’extrémité du cordond’infection. La différenciation en bactéroïdes commencealors. EPB, espace péribactéroïdien ; MS, membrane dusymbiosome ; RE, réticulum endoplasmique. D’aprèsPatriarca et al., 2004.

9
les Rhizobia. De la périphérie versl’intérieur du nodule, on trouve :• le cortex externe constitué en
majorité par des cellulesparenchymateuses
• le cortex moyen• les tissus vasculaires constitués
surtout de phloème et entourés par unendoderme et un péricycle
• le cortex interne formé de une àtrois couches de cellules
• le parenchyme central qui contientles cellules infectées par les Rhizobiaet des cellules non infectées pluspetites.
III. Les symbiosesactinorhiziennes
1) GénéralitésLes plantes actinorhiziennes, appelées
ainsi car elles forment une symbiosefixatrice d’azote avec un actinomycète,Frankia, sont pour la plupart des espècesligneuses. Elles comprennent des plantesdistribuées en 8 familles d’angiospermeset 24 genres, parmi lesquelles on trouve lefilao (Casuarina equisetifolia), l’aulne(Alnus sp.), l’olivier de Bohème (Elaeagnusangustifolia) et le myrte des marais (Myricagale). Ce sont essentiellement des arbresou arbustes adaptés aux stressédaphiques comme la salinité élevée, lesmétaux lourds ou les pH extrêmes(Dawson, 1990). Leur importance résultesurtout du fait qu’elles sont responsablede la moitié de la fixation biologique del’azote et que ce sont des espècescapables de réhabiliter des sites dégradésou de protéger les sols contre diversesformes d’érosion (Moiroud, 1996).
2) Aspects phylogénétiques etévolutifsLa répartition phylogénétique des
plantes actinorhiziennes semble assezdisparate. La capacité à former unesymbiose avec Frankia ne se retrouve pasforcément au niveau d’une même famille,ni même d’un genre. Par exemple, Dryasdrumondii est la seule espèceactinorhizienne de ce genre. Dans lafamille des Bétulacées, des genres très
proches comme Alnus et Betula nepartagent pas la capacité à fixer de l’azote(Huss-Danel, 1997). Une étudephylogénétique des plantesactinorhiziennes basée sur l’analyse dugène rbcL a permis de les grouper dans unmême clade, Rosid I, qui inclut lesLégumineuses et d’autres plantes nonsymbiotiques. Cela suggère l’existenced’une prédisposition à la symbiose au seinde ce groupe (Soltis et al, 1995).L’association avec Frankia se seraitproduite indépendamment au moinsquatre fois au cours de l’évolution(Swensen, 1996).
Les traces les plus anciennes de plantesappartenant à des familles où l'on trouveactuellement des plantes actinorhiziennessont des grains de pollen de Myricacées etd’Alnus datant de la fin du crétacé (87,5-84 millions d’années) ou du début dutertiaire (87,5-66 millions d’années). Dupollen caractéristique des Casuarinacéesest abondant pendant le paléocène (66-58millions d’années). Pour les Datiscacées,des mégafossiles correspondent à 55-39millions d’années et pour les autresfamilles de plantes actinorhiziennes, lestraces fossiles sont plus récentes : 39-23,5 millions d’années pour lesElaeagnacées, les Rhamnacées et lesRosacées, 11-5 millions d’années pour lesCoriariacées. Il n'y a pas de preuve que cesplantes aient établi des symbiosesactinorhiziennes dès le début, maiscomme les fossiles retrouvéscorrespondent dans la plupart des cas àdes milieux aux conditions nutritionnellesdifficiles comme ceux colonisés par lesplantes actinorhiziennes actuelles, cettehypothèse semble probable (Benson etClawson, 2000). Après les glaciations, lesplantes actinorhiziennes ont pu jouer unrôle écologique considérable en colonisantles moraines et les terres récemmentmises à nu par la fonte des glaciers(Benson et Clawson, 2000).
3) Le partenaire bactérienFrankia est une bactérie filamenteuse
Gram positive (Benson et Silvester, 1993).Il s’agit plus particulièrement d’unactinomycète en raison de ses

10
caractéristiques morphologiques etbiochimiques. Les colonies de Frankiacultivées in vitro se développent sous laforme de filaments et de manière radiale(Figure 10). On distingue des structuresspécialisées (hyphes végétatives septées,sporanges et vésicules). Au plan
biochimique, Frankia s’apparente auxautres actinomycètes en raison de lateneur élevée en guanine et cytosine deson ADN. Le genre Frankia constitue ungroupe homogène et se distingue par unfaisceau de caractères des autres genresd’actinomycètes. Parmi ces caractères, ilfaut noter un pouvoir infectif vis-à-vis desplantes actinorhiziennes. Il existe unespécificité pour l’infectivité (aptitude ànoduler) des souches de Frankia (Mullin etDobritsa, 1996).
L’interaction entre Frankia et les plantesactinorhiziennes conduit à la formationd’un nouvel organe racinaire appelénodule actinorhizien ou actinorhize danslequel la bactérie est hébergée et fournitde l’azote réduit à la plante.
4) La formation du noduleactinorhizienL’émergence de deux espèces modèles
chez les Légumineuses, M. truncatula et L.japonicus, a contribué à accélérer lacaractérisation génétique de la symbioseRhizobium-Légumineuse. Dans le cas dessymbioses actinorhiziennes, aucune plantemodèle n’a encore émergé. Toutes lesplantes actinorhiziennes, à l’exception deDatisca glomerata, sont des plantespérennes ligneuses ce qui rend lesapproches génétiques difficiles pour ne
pas dire impossibles. De plus, latransformation génétique de Frankia n’estpas maîtrisée ce qui limite les analyses desgènes symbiotiques bactériens. Pourtoutes ces raisons, les mécanismesmoléculaires de la formation du noduleactinorhizien sont encore largementméconnus.
Les gènes de plantes exprimés demanière spécifique en réponse à l’infectionpar Frankia sont appelés gènes denodulines actinorhiziennes par homologieaux nodulines des Légumineuses. Ondistingue les nodulines précoces,exprimées pendant les premières phasesde l’infection, et les nodulines tardivesimpliquées dans le fonctionnement dunodule. Le récent séquençage du génomede trois souches de Frankia (Normand etal, 2006) associé au développementd’approches d’ARN interférent pour desplantes actinorhiziennes commeAllocasuarina verticillata et Casuarinaglauca (C. Franche, notre laboratoire)ouvrent de nouvelles perspectives pouridentifier des gènes bactériens et végétauximpliqués dans l’interaction symbiotique.
a) PréinfectionLe développement du nodule
actinorhizien ne se produit qu’en situationde carence azotée. Les racines émettentdes signaux de nature inconnue qui sontperçus par Frankia. Les phénylpropanoïdessont suspectés jouer ce rôle par analogieavec les Légumineuses. En effet, lanodulation de l’aulne (Alnus sp.) estaméliorée par l’addition de flavonones(Benoit et Berry, 1997) ou de flavonols(Hughes et al, 1999).
En réponse à ces signaux végétaux,Frankia produit un signal symbiotique quiest perçu par la racine. Chez les plantes àvoie d’infection intracellulaire, cela conduità la courbure du poil racinaire. La naturebiochimique des facteurs actinorhiziensest toujours inconnue cependant il semblequ’elle diffère de celle des facteurs Nod deRhizobium. En effet, l’application defacteurs Nod n’a aucun effet sur lesracines d’aulne (Cérémonie et al, 1999).De plus, une fraction contenant le facteurde déformation du poil racinaire a été
Figure 10. Filaments de FrankiaFrankia est une bactérie filamenteuse Gram positive.Les hyphes se développent de manière radiale sousforme de filaments. Barre : 50µm.

11
caractérisée biochimiquement démontrantla différence de nature avec les facteursNod (Cérémonie et al, 1999). L’analyse dugénôme de Frankia indique l’absence desgènes requis pour la production desfacteurs Nod (Normand et al, 2006). Enfin,l’utilisation d’ADN génomique de Frankian’a jamais permis de complémenter desmutants nod- de Rhizobium ce quisuggère que les gènes impliqués dans laproduction des facteurs actinorhiziens etdes facteurs Nod sont différents(Cérémonie et al, 1998). À ce jour, aucungène exprimé spécifiquement en réponseà ces facteurs n’a été identifié chez lesplantes actinorhiziennes. Un tel gènemarqueur permettrait de faire progresserl’identification des facteurs actinorhiziens.
La bactérie Frankia produit desphytohormones qui pourraient jouer unrôle dans la signalisation symbiotique. Desauxines naturelles comme l’acide 3-indoleacétique (IAA) ou l’acide phénylacétique(PAA) sont présentes dans le milieu deculture de certaines souches de Frankia àdes concentrations assez élevées pour êtreperçues par la plante (10-5 à 10-6M ;Wheeler et al, 1984 ; Hammad et al, 2003).La cytokinine isopentényl adénosine (iPA) aégalement été détectée à desconcentrations de l’ordre de 10-6M(Gordons et al, 1988). Cependant, le rôleexact de ces hormones dans le processusde nodulation est toujours inconnu.
b) InfectionIl existe deux modes d’infection des
plantes actinorhiziennes par Frankia : lavoie intracellulaire et la voie intercellulaire(Figure 11). Le type d’infection estdépendant de la plante hôte (Berry etSunnel, 1990).
L’infection intracellulaire commenceavec la déformation des poils racinairesenviron 24 à 48 heures après inoculation(Figure 12A). Suivant les espèces, tous lespoils racinaires (par ex. Casuarina ; Torrey,1976) ou seulement quelques-uns (parex. Comptonia ; Callaham et al, 1979) sedéforment. Seuls les jeunes poilsracinaires en croissance et pas encorecomplètement différenciés sontcompétents pour l’infection (Callaham et
Torrey, 1977 ; Callaham et al, 1979). Leshyphes de Frankia sont piégés dans despolysaccharides d’origine végétale àl’extrémité du poil racinaire etcommencent à former un cordond’infection (Berry et al, 1986 ; Berg,1999a).
Une désorganisation locale de la paroiest observée au site de pénétration deshyphes (Berry et al, 1986). Tout au long del’infection, les hyphes restent entouréespar la membrane plasmique de l’hôte. Ungène de Casuarina glauca, nommé Cg12,codant pour une protéase de typesubtilisine est exprimé de manièrespécifique dans les cellules infectées et
Figure 11. Infections intra et inter-cellulaire chez les plantesactinorhiziennesInfection intracellulaire (Myrica, Comptonia, Alnus etCasuarina)La voie de pénétration de Frankia (Fr) est un poil absorbant. Demanière concomitante à la progression de Frankia dans le poilabsorbant, a lieu une activité mitotique dans le cortex de laracine. Les divisions cellulaires et l’hypertrophie de ces cellulesinfectées par Frankia donnent naissance à une légèreprotubérance de la racine, visible à l’œil nu, le prénodule (P). Lelobe nodulaire (LN) est initié dans des cellules du péricycle quisont opposées à l’un des pôles du xylème primaire. L’invasiondu lobe par Frankia se réalise par des hyphes issus duprénodule.Infection intercellulaire (Elaeagnus, Ceanothus et Cercocarpus)La pénétration des hyphes de Frankia a lieu dans la lamellemoyenne de deux cellules du rhizoderme. L’infection de laracine se poursuit par la progression intercellulaire des hyphesvers le lobe nodulaire qui est issu de cellules du péricycleopposées au pôle du protoxylème. L’invasion du lobe nodulairepar Frankia est intracellulaire et reste localisée au parenchymecortical du lobe nodulaire. Dans ce type d’infection, onn’observe pas de stade prénodule. D’après Franche et al., 1998.

12
notamment les poils racinaires (Laplazeet al, 2000b ; Svistoonoff et al, 2003). Ils’agit d’une protéine sécrétée qui pourraitêtre impliquée dans le remodelage de laparoi au cours de l’infection par Frankia.Les poils racinaires infectés ont une forteactivité métabolique alors que les poilsnon infectés dégénèrent rapidement (Berryet al, 1986 ; Berg, 1999a). L’infectionintracellulaire provoque la division de
quelques cellules corticales situées àproximité ce qui entraîne l’apparitiond’une protubérance à la surface de laracine que l’on appelle le « prénodule »(Figure 12B ; Callaham et Torrey, 1977). Laformation du prénodule est une étapeobligatoire avant la formation du nodulemais le prénodule n’est pas directementimpliqué dans la formation du nodule(Laplaze et al, 2000a). Les hyphes de
Figure 12. Processus d’infection intracellulaire et organogenèse d’un lobe nodulaire chez les plantes actinorhiziennesL’échange de signaux entre Frankia et la plante actinorhizienne aboutit à la déformation du poil racinaire (A). La bactérie pénètre dans un poildéformé et provoque des divisions des cellules corticales (B). Les cellules corticales en division sont infectées par les hyphes de Frankia ets’hypertrophient ce qui provoque l’apparition du prénodule. En même temps, quelques cellules du péricycle situées en face du pôle dexylème se divisent pour donner naissance au primordium nodulaire (C). Les hyphes de Frankia en provenance du prénodule envahissent lescellules corticales du nodule en formation (D). Dans le nodule mature, quatre zones sont décrites : la zone méristématique toujoursdépourvue de Frankia, la zone d’infection qui contient les cellules corticales nouvellement produites en cours d’infection, la zone defixation où les cellules infectées fixent activement l’azote atmosphérique et la zone de sénescence chez les nodules âgés (non représentée)où Frankia et les cellules végétales dégénèrent.

13
Frankia progressent de manièreintracellulaire du site d’infection endirection du prénodule au travers de pontscytoplasmiques appelés cordons depréinfection (Berg, 1999b) et qui sontformés avant infection par Frankia par unsignal de nature inconnue. Les cellules duprénodule sont infectées par Frankia et sedifférencient pour fixer l’azoteatmosphérique (Angulo Carmona, 1974 ;Laplaze et al, 2000a).
L’infection intercellulaire débute par lapénétration de Frankia entre les cellules del’épiderme (Figure 12). Aucunedéformation des poils racinaires n’estassociée à ce type d’infection. Un matériaudense est sécrété par les cellules végétalesau site d’infection (Miller et Baker, 1985).Frankia progresse dans les tissus corticauxau travers de la lamelle moyenne.
c) Développement du noduleAu cours des deux types d’infection
(voie intracellulaire ou intercellulaire),Frankia induit la division de certainescellules du péricycle situées en face dupôle de xylème pour donner naissance àun primordium nodulaire (Figure 12C etD). Chez Comptonia, certaines cellulescorticales subissent des divisions etparticipent à la formation du primordium(Callaham et Torrey, 1977). Chaque lobenodulaire présente un méristème apical,un système vasculaire central et unpériderme à sa périphérie. Les hyphes deFrankia soit en provenance du prénodule(infection intracellulaire) soit enprogression intercellulaire à travers lecortex (infection intercellulaire)envahissent les cellules corticales situées àla base du primordium nodulaire. Laprogression de Frankia à l’intérieur dunodule se fait en direction de l’apex ce quicrée un gradient de développementcentrifuge de Frankia et des cellulesinfectées (Figure 12E).
d) Structure du noduleQuatre zones sont habituellement
décrites :• la zone méristématique située à
l’apex. Elle est responsable de lacroissance indéterminée du nodule.
Cette zone est toujours dépourvue deFrankia.
• la zone d’infection adjacente auméristème apical. Les hyphes infectentles cellules corticales nouvellementformées.
• la zone de fixation qui contient à lafois des cellules infectées et descellules non infectées. Les cellulesinfectées présentent une hypertrophiecaractéristique et sont rempliesd’hyphes. Les cellules bactériennes etvégétales se différencient pourpermettre la fixation de l’azote.
• la zone de sénescence qui estprésente chez les nodules âgés. Lescellules bactériennes et végétalesdégénèrent, Frankia produit dessporanges.
Chez certaines espèces commeCasuarina ou Myrica, une « racinenodulaire » dépourvue de Frankia estprésente à l’apex de chaque lobenodulaire (Figure 13). C’est une racine
modifiée qui présente un gravitropismenégatif (elle pousse vers la surface). Ellecomporte un aérenchyme développé cequi laisse supposer qu’elle jouerait un rôledans la circulation des gaz (Pawlowski etBisseling, 1996). Dans ce cas, leméristème nodulaire est reprogrammépour devenir un méristème de racinenodulaire mais les mécanismesmoléculaires responsables de cechangement sont inconnus.
Figure 13. Nodule et racines nodulaires de CasuarinaglaucaPhotographie d’un nodule âgé de 5 semaines, présentantplusieurs lobes qui comportent une racine nodulaire à l’apex.Barre : 500 µm. D’après Laplaze, 1999.

14
La croissance du nodule se fait parramification des lobes nodulaires. Unnouveau primordium est formé à partirdes cellules du péricycle du lobepréexistant puis il est infecté par leshyphes de Frankia en provenance dunodule. Ce mode de croissance déterminela structure coralloïde des nodulesactinorhiziens (Figure 13).
e) Le nodule actinorhizien : une racinelatérale modifiée ?
Du fait de son originedéveloppementale identique à celle d’uneracine latérale (division des cellules dupéricycle situées en face du pôle dexylème) et de son anatomie similaire àcelle d’une racine latérale, le noduleactinorhizien est souvent considérécomme une racine latérale modifiée(Pawlowski et Bisseling, 1996). Laprésence de la racine nodulaire à l’apex dunodule chez certaines espèces de plantesactinorhiziennes est en accord avec cettethéorie. Cependant, une étudeapprofondie menée chez Alnus glutinosasuggère que les nodules ne sont pasformés à partir de racines latéralespréexistantes (Angulo Carmona, 1974). Deplus, les cellules corticales peuvent êtreimpliquées dans la formation duprimordium nodulaire chez certainesespèces comme Comptonia alors que leprimordium de racine latérale provientuniquement des cellules du péricycle(Callaham et Torrey, 1977). Enfin, bien quele nodule présente les caractéristiquesdéveloppementales et morphologiquesd’une racine latérale, l’existence de voiesde développement communes entre cesdeux organes n’a jamais été démontrée.
L’infection par Frankia ne modifie pas ledéveloppement du système racinairemême si les plantes nodulées ont unemeilleure croissance que les plantes noninoculées. L’étroite ressemblance entre lesnodules actinorhiziens et les racineslatérales aurait pu laisser penser quel’infection a un impact sur l’architectureracinaire. Une étude fine de la nodulationd’Alnus glutinosa a permis de montrer quela nodulation n’a aucun impact sur ladistribution des racines latérales (Angulo
Carmona, 1974). Les nodulesactinorhiziens sont donc des organessupplémentaires qui ne se forment pas àpartir de primordia de racines latéralespréexistants. De plus, il n’y a pasd’augmentation du nombre de racineslatérales en réponse à l’infection (AnguloCarmona, 1974). Ces résultats suggèrentque la formation du nodule actinorhizienet de la racine latérale sont régulés demanière indépendante. Néanmoins, ledéveloppement de la racine latérale est unbon modèle pour l’analyse dudéveloppement du nodule actinorhiziendu fait de la grande ressemblanceanatomique entre ces deux organes.
IV. Formation de la racinelatérale
1) GénéralitésAprès germination, les parties
aériennes produisent des méristèmes auxaisselles de chaque primordium foliaire.Ces méristèmes secondaires sontconstitués de cellules indifférenciées quipeuvent être activées pour permettre laramification du système aérien. À l’inverse,aucun méristème secondaire n’est générépar le méristème apical racinaire. De cefait, la ramification du système racinaireimplique la formation de novo deméristèmes à partir de cellulesdifférenciées (Dolan et Scheres, 1998).Cependant, la détermination des cellules àl’origine des primordia de racine latéralepourrait se faire très tôt au cours de lamise en place de la racine primaire (De
Figure 14. Différents stades de formation duprimordium de racine latérale chez ArabidopsisthalianaLes cellules du péricycle donnent naissance au primordiumde racine latérale. La mise en place du primordium est unmécanisme très régulé et bien décrit. IL, inner layer ; OL,outer layer. D’après Malamy et Benfey, 1997.

15
Smet et al, 2007).L’organisation très simple de la racine
d’Arabidopsis a permis une caractérisationhistologique fine des mécanismes de miseen place du méristème de racine latérale(Figure 14 ; Malamy et Benfey, 1997). Ladisponibilité de nombreux mutants aconduit à l’identification de nombreuxgènes impliqués dans la formation de laracine latérale (Casimiro et al, 2003). Lamise en place de la racine se découpe entrois étapes majeures : initiation,organisation du primordium etémergence.
2) InitiationLa racine d’Arabidopsis a une anatomie
assez simple puisqu’elle est composéed’une série de couches monocellulairesautour des tissus vasculaires (xylème etphloème) : le péricycle, l’endoderme, lecortex et l’épiderme (Figure 15 ; Dolan et
al, 1993). Les racines latérales proviennentdes cellules du péricycle situées en facedes deux pôles de xylème (Dubrovsky etal, 2001). La plupart des cellules dupéricycle sont bloquées en phase G1 etseules les cellules compétentes, situées enface des pôles de xylème, progressent
vers la phase S puis G2 (Figure 16 ;Beeckman et al, 2001).
Parmi les cellules compétentes, lescellules « fondatrices » sont définiescomme des cellules qui acquièrent undestin développemental différent desautres cellules du péricycle et qui, parconséquent, jouent un rôle primordialdans l’initiation de la racine latérale(Dubrovsky et al, 2001). Les premiersévénements morphologiques associés àl’initiation d’une racine latérale ont lieudans deux cellules fondatrices adjacentes(De Smet et al, 2006). Ces cellules sedivisent de manière anticlinale etasymétrique pour créer deux cellulescourtes flanquées de deux celluleslongues (De Smet et al, 2006). Cette
Figure 15. Anatomie de la racine d’Arabidopsis thalianaLa racine d’A. thaliana possède une organisation très simple,le tissu vasculaire est entouré de quatre couchesmonocellulaires : le péricycle, l’endoderme, le cortex etl’épiderme. D’après Benfey et Scheres, 2000.
Figure 16. Régulation du cycle cellulaire dans lepéricycleDans le méristème racinaire, les divisions cellulaires sonttrès actives (A). En amont de cette zone, les tissusexternes sortent du cycle cellulaire et se différencientalors que les tissus internes continuent leurs divisions(B). Puis les cellules du péricycle stoppent leurs divisionset restent bloquées en phase G1 (C). Les cellules dupéricycle situées en face d’un des pôles de xylèmeprogressent vers la phase S puis G2 et deviennent alorscompétentes pour l’initiation de la racine latérale (D).D’après Beeckman et al., 2001.

16
première division détermine la naissanced’un primordium de stade I (Malamy etBenfey, 1997).
L’auxine est considérée comme lesignal induisant l’initiation. En effet,l’apport d’auxine exogène induit laformation de racines latérales. De plus, laperturbation du transport polarisé del’auxine inhibe le développement desracines latérales (Reed et al, 1998). Lesanalyses génétiques ont permisd’identifier de nombreux mutants affectésdans la réponse à l’auxine (axr, tir1, shy2),son transport (aux1) et son homéostasie(alf1, ydk1, dfl1) et présentant unphénotype au niveau des racines latérales(De Smet et al, 2006). Notamment, letransport d’influx d’auxine est impliquédans l’étape d’initiation puisque le mutantaux1 présente une réduction de 50% deprimordia initiés (Marchant et al, 2002).Chez le mutant aux1, les niveaux d’auxinesont suboptimaux pour permettrel’initiation du primordium mais les étapessuivantes de mise en place du méristèmene semblent pas affectées.
Les racines latérales sont produites àdistance du méristème apical et de la zoned’élongation (Dubrovsky et al, 2000).Cependant, des études récentes montrentque les cellules fondatrices pourraient êtredéterminées dans le méristème basal sousl’action de l’auxine (De Smet et al, 2007).Le méristème basal correspond à la régionsituée entre le méristème apical et la zoned’élongation (Beemster et al, 2003) etprésente une sensibilité élevée à l’auxine.Dans cette zone, les pics de perception del’auxine correspondent aux futurs sitesd’initiation de la racine latérale. Un signalauxinique récurrent perçu par leméristème basal permettrait donc dedéterminer la distribution des racineslatérales sur l’axe racinaire (De Smet et al,2007).
L’utilisation du marqueur de perceptionde l’auxine DR5:GUS montre que l’auxines’accumule dans les cellules du péricycleavant et pendant les premières divisionsasymétriques (Benková et al, 2003). Lemutant slr1 (solitary root) est incapabled’initier des racines latérales (Fukaki et al,2002). Chez ce mutant, la protéine IAA14
(Aux/IAA) est stabilisée et inhibe la voie designalisation auxinique qui mènenormalement à l’initiation. L’interactioninhibitrice implique les protéines ARF7 etARF19 et a pour effet d’empêcherl’activation des cellules du péricycle et lespremières divisions anticlinales (Fukaki etal, 2005 ; Okushima et al, 2007).
Les cibles de la signalisation auxiniquedans les cellules du péricycle ne sont pastoutes identifiées. Cependant, il est clairque l’auxine est le signal qui permetl’activation des cellules du péricycle enleur faisant passer la transition G1-Sd’entrée dans le cycle cellulaire (Himanenet al, 2002 ; Vanneste et al, 2005b). Legène KRP2, un inhibiteur des kinasesdépendantes de cycline (CDK) et donc dela transition G1-S, est exprimé dans lescellules du péricycle qui ne sont pas àl’origine des racines latérales (Himanen etal, 2002). La surexpression de KRP2entraine une diminution de l’ordre de 60%du nombre de racines latérales.L’activation des cellules du péricycle pourla formation des racines latérales pourraitdonc passer par une levée d’inhibition deKRP2. En effet, l’expression du gène KRP2est inhibée par l’auxine (Himanen et al,2002) ce qui suggère que l’auxine agitdirectement sur le cycle cellulaire. Pourautant, l’activation du cycle cellulaire n’estpas la seule cible de la signalétiqueauxinique car elle n’est pas suffisante pourpermettre la formation d’une racinelatérale. En effet, la surexpresion du gèneCYCD3;1 codant une cycline de type Dchez le mutant slr entraine des divisionscellulaires dans le pericycle mais nepermet pas la formation de racineslatérales (Vanneste et al, 2005a). Desanalyses transcriptomiques ont étéréalisées afin d’identifier de nouvellescibles de la signalisation auxinique(Vanneste et al, 2005b). L’étape d’initiationde la racine latérale nécessite donc quel’activation du cycle cellulaire soitaccompagnée d’une détermination descellules fondatrices. Les recherches encours devraient permettre de préciserquels sont les événements qui conduisentà cette détermination.

17
3) Organisation du primordiumLes cellules filles issues des cellules du
péricycle poursuivent leurs divisionssymétriques et asymétriques pour donnerun groupe de 8 à 10 cellules courtes detaille identique. Les cellules entamentensuite une série de divisions périclinalespour donner naissance à un primordiumcomportant deux (stade II), puis trois(stade III) et enfin quatre couchescellulaires (stade IV). À partir du stade II, leprimordium progresse à travers lapremière couche externe : l’endoderme(Malamy et Benfey, 1997). L’organisationdu primordium aboutit à une structureméristématique identique à celle duméristème primaire à partir du stade VI.L’utilisation d’un certain nombre demarqueurs spécifiques de l’endoderme,du cortex ou de l’épiderme a permis demettre en évidence la différenciation deces types cellulaires indiquant uneorganisation du primordium de racinelatérale similaire à celle de la racineprimaire (Malamy et Benfey, 1997).
L’utilisation du marqueur DR5:GUS apermis de montrer qu’un gradientd’auxine est formé dans le primordium,avec le maximum d’auxine formé à l’apex.Ce gradient est nécessaire à la formationdu méristème racinaire latéral (Benková etal, 2003 ; Swarup et Bennett, 2003) et samise en place requiert l’action coordonnéede transporteurs d’influx et de manièreprépondérante d’efflux d’auxine(Wisniewska et al, 2006). La mutation desgènes de transport d’efflux d’auxine (PIN)n’a qu’un effet limité sur l’organisation duméristème (Gälweiler et al, 1998).Néanmoins, l’utilisation de mutantsmultiples permet de contourner laredondance fonctionnelle de ces gènes etfait apparaître un rôle important dans lamise en place de ce gradient. Par exemple,chez les triples mutants pin1pin4pin7 oupin1pin3pin7, l’auxine induit la formationde primordia de racine latéraleressemblant à des massifs cellulaires peuorganisés (Benková et al, 2003). Unphénotype identique est obtenu parinhibition du transport d’auxine (Benkováet al, 2003). La régulation de la localisationcellulaire des protéines PIN est également
importante pour l’organisation duprimordium. Le gène GNOM code unfacteur d’échange pour les GTPases de lafamille des facteurs de ribosylation del’ADP (ARF-GEF) qui régule le trafficvésiculaire et est requis pour la localisationde PIN1 (Geldner et al, 2003). Un allèlefaible du mutant gnom présente lesmêmes défauts d’organisation duprimordium de racine latérale que lestriples mutants pin en réponse à l’auxine(Geldner et al, 2004).
4) ÉmergenceL’étape finale (stade VIII ou émergence)
correspond à la sortie du primordium de laracine principale. L’émergence n’est pasassociée avec des divisions cellulaires maisà un allongement des cellules qui permetun accroissement de la taille duprimordium. Les cellules des tissusexternes s’écartent pour laisser passer leprimordium en croissance. Aprèsémergence, les divisions cellulairesreprennent à l’apex de la racine latéralenouvellement formée (Malamy et Benfey,1997).
Les mécanismes de régulation del’étape d’émergence de la racine latéraleont été peu étudiés. Différentes approchesont permis d’identifier des gènesimpliqués dans ce processus (Neuteboomet al, 1999a ; Laskowski et al, 2006). Ilsemblerait que l’induction par l’auxine degènes de remodelage de la paroi permettele passage du primordium à travers lestissus externes. Parmi ces gènes, onretrouve notamment AtAIR3 qui estexprimé spécifiquement dans les cellulessituées en face du primordium enformation (Neuteboom et al, 1999b). Legène AIR3 code une protéase à sérine detype subtilisine qui est excrétée etpermettrait de modifier les propriétés de laparoi végétale.
En conclusion, l’auxine apparaît commele signal clé de la mise en place desracines latérales (Casimiro et al, 2001 ;Bhalerao et al, 2002). L’auxine intervientau cours des trois étapes majeures de laformation des racines latérales :l’initiation, l’organisation du méristème et

18
très probablement aussi au cours del’émergence.
V. L’hormone végétaleauxine
1) IntroductionL’auxine joue un rôle crucial dans de
nombreux mécanismes physiologiques etdéveloppementaux : développement del’embryon et du fruit, organogenèse,différenciation du système vasculaire, miseen place de l’architecture racinaire,élongation cellulaire, tropismes etdominance apicale (Figure 17 ; Kepinski etLeyser, 2005a). De nombreuses molécules
possèdent un pouvoir auxinique mais laplus abondante chez les plantes est l’acide3-indole acétique (IAA), les différentesformes naturelles et synthétiques utiliséesau cours de cette thèse sont présentéesdans la figure 18.
2) Perception et signalisationLa perception et la transduction du
signal auxinique ont fait l’objet derecherches intensives et de nombreusesrevues récentes sont disponibles (Leyser,2006 ; Paciorek et Friml, 2006 ; Quint etGray, 2006). Des récepteurs de l’auxineont récemment été identifiés, il s’agit de laprotéine TIR1 (Dharmasiri et al, 2005a ;
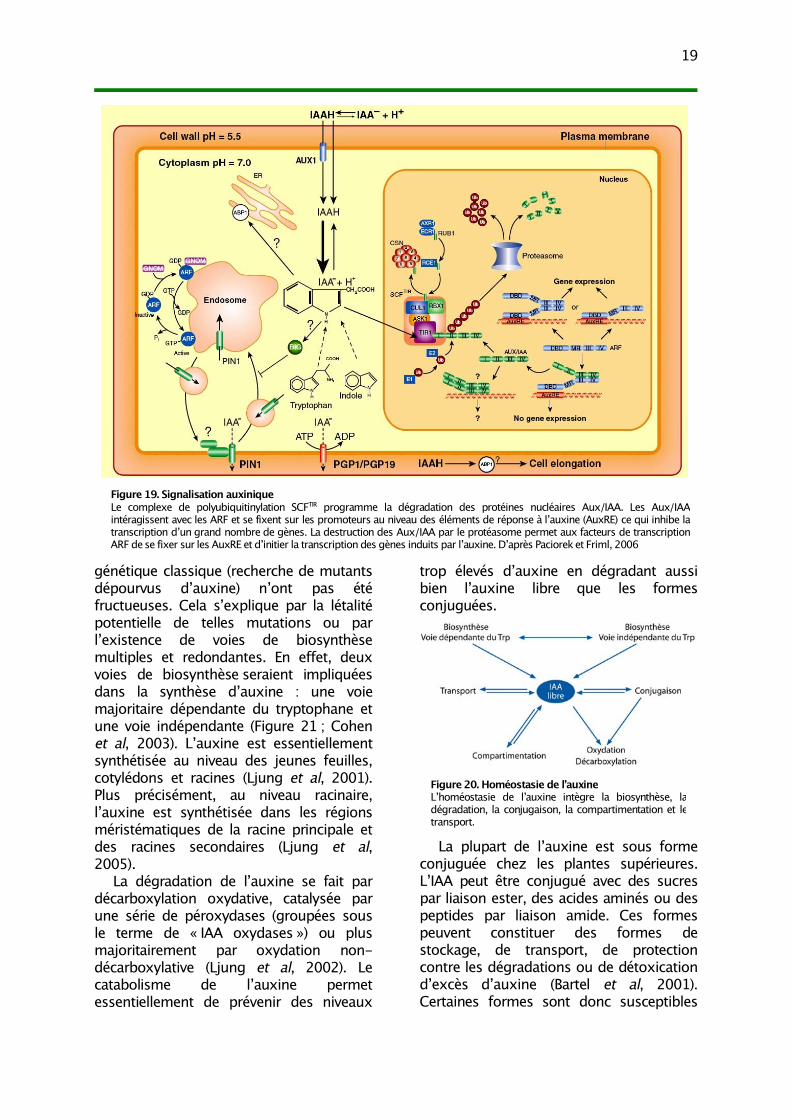
Kepinski et Leyser, 2005b) et desmembres de la famille AFB (Dharmasiri etal, 2005b) même si l’existence d’autresrécepteurs reste envisagée (Badescu etNapier, 2006). La figure 19 synthétise lasignalisation auxinique qui fait intervenirle complexe de polyubiquitinylation SCFTIR
qui programme la dégradation desprotéines Aux/IAA. Les Aux/IAAintéragissent avec les facteurs detranscription ARF et empêchent latranscription des gènes contenant deséléments AuxRE dans leur promoteur. Ladestruction des Aux/IAA par leprotéasome permet aux ARF de se fixersur les AuxRE et d’initier la transcriptiondes gènes induits par l’auxine. Cette voiede signalisation par levée d’inhibition peutsembler complexe mais elle présentecertains avantages, notamment laprésence de nombreux Aux/IAA et ARFassure la spécificité de la réponseauxinique en fonction des types cellulaires(Teale et al, 2006).
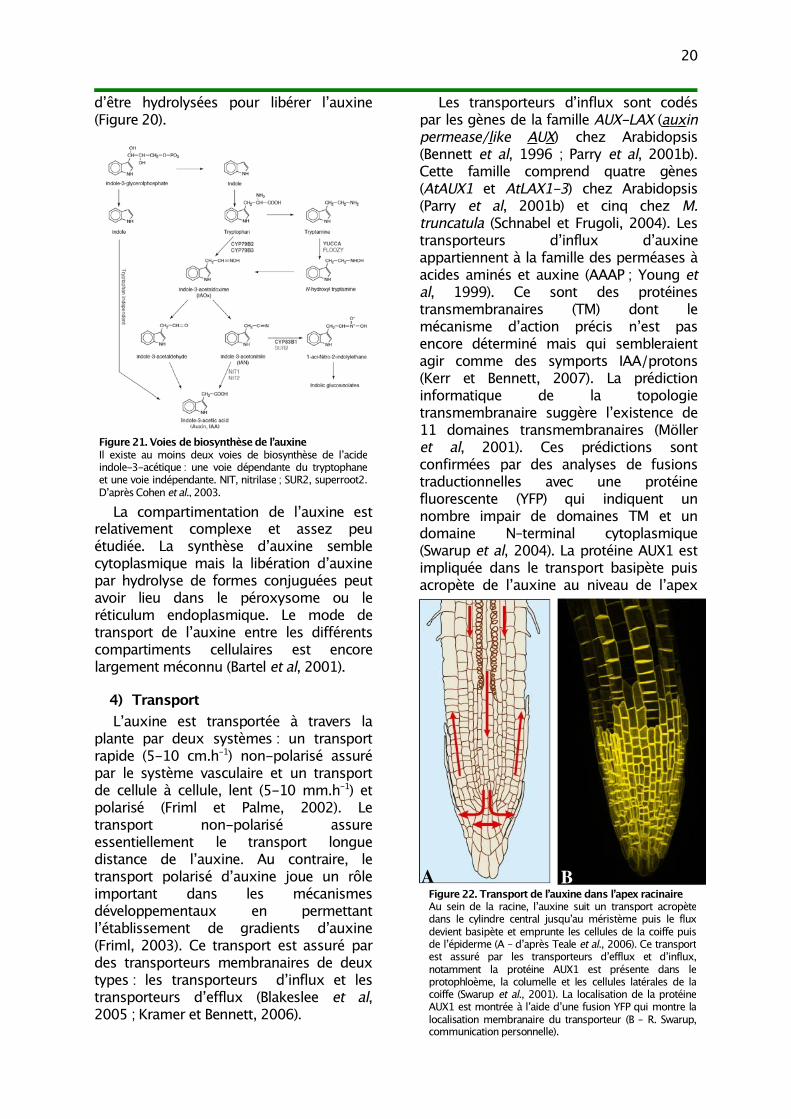
3) HoméostasieL’homéostasie de l’auxine est complexe
puisqu’elle intègre la biosynthèse, ladégradation, la conjugaison, lacompartimentation et le transport (Figure20 ; Ljung et al, 2002 ; Woodward etBartel, 2005).
Les détails de la biosynthèse de l’auxinene sont pas totalement connus (Woodwardet Bartel, 2005). Les approches de
Figure 17. Implication des flux d’auxine dans lesprocessus développementauxLe transport d’auxine est impliqué dans de nombreuxmécanismes développementaux chez les plantes :rhizogénèse latérale (a), organisation de l’embryon (b),phyllotaxie (c), différenciation du tissu vasculaire foliaire(d) et organisation du méristème apical racinaire (e).D’après Teale et al., 2006.
Figure 18. Formules chimiques des différents typesd’auxineLa formule chimique des principaux types d’auxinenaturels (IAA, IBA et PAA) et synthétiques (2,4D et NAA)est présentée ainsi que l’inhibiteur compétitif dutransport d’influx (NOA).
19
génétique classique (recherche de mutantsdépourvus d’auxine) n’ont pas étéfructueuses. Cela s’explique par la létalitépotentielle de telles mutations ou parl’existence de voies de biosynthèsemultiples et redondantes. En effet, deuxvoies de biosynthèse seraient impliquéesdans la synthèse d’auxine : une voiemajoritaire dépendante du tryptophane etune voie indépendante (Figure 21 ; Cohenet al, 2003). L’auxine est essentiellementsynthétisée au niveau des jeunes feuilles,cotylédons et racines (Ljung et al, 2001).Plus précisément, au niveau racinaire,l’auxine est synthétisée dans les régionsméristématiques de la racine principale etdes racines secondaires (Ljung et al,2005).
La dégradation de l’auxine se fait pardécarboxylation oxydative, catalysée parune série de péroxydases (groupées sousle terme de « IAA oxydases ») ou plusmajoritairement par oxydation non-décarboxylative (Ljung et al, 2002). Lecatabolisme de l’auxine permetessentiellement de prévenir des niveaux
trop élevés d’auxine en dégradant aussibien l’auxine libre que les formesconjuguées.
La plupart de l’auxine est sous formeconjuguée chez les plantes supérieures.L’IAA peut être conjugué avec des sucrespar liaison ester, des acides aminés ou despeptides par liaison amide. Ces formespeuvent constituer des formes destockage, de transport, de protectioncontre les dégradations ou de détoxicationd’excès d’auxine (Bartel et al, 2001).Certaines formes sont donc susceptibles
Figure 19. Signalisation auxiniqueLe complexe de polyubiquitinylation SCFTIR programme la dégradation des protéines nucléaires Aux/IAA. Les Aux/IAAintéragissent avec les ARF et se fixent sur les promoteurs au niveau des éléments de réponse à l’auxine (AuxRE) ce qui inhibe latranscription d’un grand nombre de gènes. La destruction des Aux/IAA par le protéasome permet aux facteurs de transcriptionARF de se fixer sur les AuxRE et d’initier la transcription des gènes induits par l’auxine. D’après Paciorek et Friml, 2006
Figure 20. Homéostasie de l’auxineL’homéostasie de l’auxine intègre la biosynthèse, ladégradation, la conjugaison, la compartimentation et letransport.
20
d’être hydrolysées pour libérer l’auxine(Figure 20).
La compartimentation de l’auxine estrelativement complexe et assez peuétudiée. La synthèse d’auxine semblecytoplasmique mais la libération d’auxinepar hydrolyse de formes conjuguées peutavoir lieu dans le péroxysome ou leréticulum endoplasmique. Le mode detransport de l’auxine entre les différentscompartiments cellulaires est encorelargement méconnu (Bartel et al, 2001).
4) TransportL’auxine est transportée à travers la
plante par deux systèmes : un transportrapide (5-10 cm.h-1) non-polarisé assurépar le système vasculaire et un transportde cellule à cellule, lent (5-10 mm.h-1) etpolarisé (Friml et Palme, 2002). Letransport non-polarisé assureessentiellement le transport longuedistance de l’auxine. Au contraire, letransport polarisé d’auxine joue un rôleimportant dans les mécanismesdéveloppementaux en permettantl’établissement de gradients d’auxine(Friml, 2003). Ce transport est assuré pardes transporteurs membranaires de deuxtypes : les transporteurs d’influx et lestransporteurs d’efflux (Blakeslee et al,2005 ; Kramer et Bennett, 2006).
Les transporteurs d’influx sont codéspar les gènes de la famille AUX-LAX ( aux inpermease/ l ike A U X ) chez Arabidopsis(Bennett et al, 1996 ; Parry et al, 2001b).Cette famille comprend quatre gènes(AtAUX1 et AtLAX1-3) chez Arabidopsis(Parry et al, 2001b) et cinq chez M.truncatula (Schnabel et Frugoli, 2004). Lestransporteurs d’influx d’auxineappartiennent à la famille des perméases àacides aminés et auxine (AAAP ; Young etal, 1999). Ce sont des protéinestransmembranaires (TM) dont lemécanisme d’action précis n’est pasencore déterminé mais qui sembleraientagir comme des symports IAA/protons(Kerr et Bennett, 2007). La prédictioninformatique de la topologietransmembranaire suggère l’existence de11 domaines transmembranaires (Mölleret al, 2001). Ces prédictions sontconfirmées par des analyses de fusionstraductionnelles avec une protéinefluorescente (YFP) qui indiquent unnombre impair de domaines TM et undomaine N–terminal cytoplasmique(Swarup et al, 2004). La protéine AUX1 estimpliquée dans le transport basipète puisacropète de l’auxine au niveau de l’apex
Figure 21. Voies de biosynthèse de l’auxineIl existe au moins deux voies de biosynthèse de l’acideindole-3-acétique : une voie dépendante du tryptophaneet une voie indépendante. NIT, nitrilase ; SUR2, superroot2.D’après Cohen et al., 2003.
A BFigure 22. Transport de l’auxine dans l’apex racinaireAu sein de la racine, l’auxine suit un transport acropètedans le cylindre central jusqu’au méristème puis le fluxdevient basipète et emprunte les cellules de la coiffe puisde l’épiderme (A – d’après Teale et al., 2006). Ce transportest assuré par les transporteurs d’efflux et d’influx,notamment la protéine AUX1 est présente dans leprotophloème, la columelle et les cellules latérales de lacoiffe (Swarup et al., 2001). La localisation de la protéineAUX1 est montrée à l’aide d’une fusion YFP qui montre lalocalisation membranaire du transporteur (B – R. Swarup,communication personnelle).

21
racinaire (Figure 22 ; Swarup et al, 2001).Plusieurs gènes pourraient coder les
transporteurs d’efflux. Les gènes de lafamille PIN (pin-formed) sont au nombrede huit chez Arabidopsis (AtPIN1-8 ;Boutté et al, 2006) et au moins cinq chezM. truncatula (Paponov et al, 2005). Ils’agit de protéines transmembranaires quipourraient assumer le rôle detransporteurs d’efflux (Petrasek et al,2006) au sein d’un complexemultiprotéique. La polarisation cellulairedes protéines PIN est responsable dutransport polarisé de l’auxine (Wisniewskaet al, 2006). Il existe une forte redondancefonctionnelle des portéines PIN associée àune régulation croisée de leurs profilsd’expression (Vieten et al, 2005). L’auxinepeut réguler l’expression des PIN àdifférents niveaux : par exemplel’expression du gène PIN7 est induite parl’auxine alors que la stabilité de la protéinePIN7 est réduite par de fortesconcentrations en auxine mais augmentéepar de faibles concentrations (Vieten et al,2005). Cette régulation complexe desprotéines PIN permet une canalisation duflux d’auxine au cours des des différentsprocessus développementaux (Sauer et al,2006).
Certaines protéines codées par lesgènes de la famille MDR/PGP ( m ulti d rugr esistance/ P - g lyco p roteins) ont égalementprouvé leur capacité à transporter l’auxinehors des cellules. Les protéines MDR/PGPappartiennent à la classe destransporteurs ABC ( A TB b inding c assette).La famille MDR/PGP comprend 21 gèneschez Arabidopsis (Martinoia et al, 2002 ;Jasinski et al, 2003) dont seulementcertains ont été identifiés comme destransporteurs d’efflux d’auxine : AtPGP1(Geisler et al, 2005), AtPGP4 (Terasaka etal, 2005) et AtPGP19 (Noh et al, 2001).
VI. Objectifs de la thèseLa famille des Casuarinacées comprend
90 espèces groupées en 4 genres :Casuarina, Allocasuarina, Gymnostoma etCeuthostoma. Plus des deux tiers de cesespèces sont originaires d’Australie, dediverses îles du Pacifique et de différentesrégions du sud-est de l’Asie
(Dommergues et al, 1999). Le nomCasuarina fait allusion aux rameaux trèsfins et tombants qui ressemblent auplumage du casoar (Casuarius casuarius).Leur système racinaire présente unegrande plasticité de développement. Eneffet, ces plantes peuvent former dessymbioses avec des champignons (ecto-ou endomycorhizes, voire les deux chezcertaines espèces comme Casuarinaglauca). En l’absence de champignonsmycorhiziens et en conditions de carenceen fer et en phosphate, des racinesprotéoïdes (cluster roots) peuventapparaître. Ce sont des touffes de racinesà croissance déterminée. Il a été montréque ces structures augmententl'acquisition de fer et de phosphate(Neumann et Martinoia, 2002). Enfin, enconditions de carence azotée, lesCasuarinacées peuvent former desnodules fixateurs d'azote (actinorhizes) ensymbiose avec l’actinomycète Frankia(Obertello et al, 2003). Ces plantes sontconsidérées comme des plantespionnières car elles colonisent les solspauvres ou dégradés permettant unerevégétalisation de ces zones(Dommergues et al, 1999).
Ce travail de thèse s’inscrit dans lecadre d’un programme de recherchevisant à étudier les mécanismesd’adaptation des Casuarinacées en sefocalisant sur la formation du noduleactinorhizien. La similitude entre ledéveloppement et l’anatomie du nodule etde la racine latérale nous a naturellementconduits à nous intéresser au rôle del’auxine. Plus précisément, nous avonsdécidé de nous focaliser sur lestransporteurs d’influx qui présententplusieurs avantages :• ils sont codés par une petite famille
multigénique chez Arabidopsis.• il n’y a pas de redondance
fonctionnelle de ces gènes chezArabidospis.
• leur fonction dans la formation de laracine est relativement bien décrite.
• la production d’auxine par Frankiasuggère que l’influx d’auxine pourraitêtre impliqué dans le processusd’infection.

22
Ce travail de thèse s’articule en troisparties :
1) Isolement et caractérisationfonctionnelle des gènes AUX-LAXchez Casuarina glaucaL’identification d’homologues des
gènes AUX-LAX a été menée paramplification d’ADN génomique à l’aided’amorces dégénérées. L’objectif était decloner tous les membres de la familleAUX-LAX chez C. glauca pour permettreune comparaison des séquences etensuite de se focaliser sur les meilleurscandidats. L’utilisation de la plante modèleArabidopsis a permis de vérifier que lesgènes identifiés chez Casuarina codentdes transporteurs d’influx d’auxine ayantune fonction équivalente. Cette analyses’est basée sur la complémentationfonctionnelle de mutants de transporteursd’influx par expression des gènes deCasuarina. L’analyse de l’expression a étémenée chez Arabidopsis afin de comparerles profils d’expression avec ceux desgènes endogènes.
2) Implication au cours dudéveloppement du noduleactinorhizienL’analyse de l’expression des gènes
AUX-LAX a été réalisée au cours de lanodulation et en réponse à différentstraitements auxiniques. Afin de confirmerle rôle du transport d’influx d’auxine etplus globalement de l’auxine au cours del’infection, l’effet d’inhibiteurs dutransport de l’auxine sur l’infection a étédéterminé. En parallèle, des dosagesd’auxine ont également été entrepris.
3) Rôle des transporteurs d’influxdans le développement : exemple deLAX3Afin de préciser le rôle du transporteur
AtLAX3 dans la mise en place de la racine,l’expression de gènes impliqués dans leremodelage de la paroi a été analysée.L’objectif était de déterminer lesmécanismes situés en aval du gèneAtLAX3 potentiellement impliqués dans lesétapes tardives de la mise en place de laracine latérale. Les résultats attendusdevaient permettre d’ouvrir des nouvellespistes de recherche sur la mise en placedu nodule actinorhizien.

23
PARTIE I - ISOLEMENT ET CARACTÉRISATION DE GÈNES AUX-LAXCHEZ CASUARINA GLAUCA
I. Isolement d’homologuesde gènes AUX-LAX chezC. glauca
1) Obtention des ADNc et desséquences génomiquesPour isoler les membres de la famille
AUX-LAX de C. glauca, une amplificationPCR sur ADN génomique a été réalisée àl’aide d’amorces dégénérées.L’amplification d’ADN génomiqueconstitue une approche sans a priori deprofil d’expression. Les amorces utiliséesont été dessinées par R. Swarup (Universitéde Nottingham, Royaume-Uni).
Le couple d’amorces (AF2 et AF3) apermis d’amplifier trois fragments d’ADNqui ont été clonés et séquencés. Lepremier fragment de 576 pdb présenteune forte homologie avec les gènes LAX3de Medicago truncatula (88% d’identité auniveau nucléique sur la partie codante dela séquence) et d’Arabidopsis thaliana(85% d’identité sur la partie codante de laséquence). La portion d’ADNccorrespondant à ce fragment génomique aété obtenue par PCR sur ADNc issus deracines, mettant en évidence un intron de128 pdb dans la séquence génomique. Leclone a été nommé CgLAX3. Le deuxièmefragment de 769 pdb présente une fortehomologie avec la plupart des membresde la famille AUX-LAX de différentesespèces (86% d’identité avec MtLAX4, 85%avec MtLAX2, 81% avec MtLAX1). Laportion d’ADNc correspondant à cefragment génomique a été obtenue,mettant en évidence un intron de 84 pdbdans la séquence génomique. Le clone aété nommé CgAUX1. Le troisièmefragment de 1200 pdb ne présenteaucune homologie avec les gènes de lafamille AUX-LAX.
Les séquences de gènes AUX-LAX desespèces suivantes ont été alignées à l’aidedu logiciel CLUSTAL-W : A. thaliana, M.truncatula, Populus tremuloides, Oryzasativa, Zea mays, Cucumis sativus. Denouvelles amorces dégénérées (AD_01,AD_02, AD_03 et AD_04) ont étédessinées à partir de ces alignements deséquences afin de tenter d’obtenir denouveaux membres de la famille AUX-LAXde C. glauca. Quatre fragments ont étéobtenus par PCR sur ADN génomique etont été séquencés. Les deux premierscorrespondent au clone CgLAX3, letroisième au clone CgAUX1 et le dernier neprésente aucune homologie avec les gènesde la famille AUX-LAX.
Pour obtenir la séquence complète desADNc correspondant à CgAUX1 et CgLAX3la technique de la RACE-PCR a été utilisée.Les amorces pour la RACE-PCR 5’ et 3’ ontété dessinées dans les exons desfragments génomiques de CgAUX1 etCgLAX3. Seul un fragment 3’ de 1236 pdba été obtenu pour CgAUX1 malgré denombreuses tentatives pour obtenir lefragment 5’. L’incapacité à obtenir cefragment 5’ par RACE-PCR nous a conduità utiliser la technique de marche sur l’ADNgénomique. Deux amplificationssuccessives ont été nécessaires pourobtenir la séquence codante complète : unpremier fragment de 832 pdb puis undeuxième de 2418 pdb ont été obtenus.Un ADNc de 1652 pdb contenant laséquence codante complète de CgAUX1 aété obtenu par amplification PCR sur ADNcà l’aide des amorces AUX1F et AUX1Rdessinées en 5’ à une cinquantaine de pdben amont de l’ATG et à l’extrémité 3’ dufragment RACE 3’. Un fragment de 3166pdb correspondant au gène CgAUX1 a étéobtenu par amplification sur ADNgénomique à l’aide des mêmes amorces.

24
Deux fragments ont pu êtreamplifiés pour CgLAX3 : un fragment 5’ de713 pdb et un fragment 3’ de 982 pdb.Ces deux fragments ont été séquencés etont permis de réaliser un contig in silicoavec la région centrale. Les amorces LAX3Fet LAX3R ont été dessinées respectivementaux extrémités 5’ et 3’ de ce contig. UnADNc de 1668 pdb contenant la séquencecodante complète de CgLAX3 a été obtenupar amplification PCR sur ADNc à l’aide deces amorces. Cette séquence comporte 3pdb en amont de l’ATG. Le gènecorrespondant (2494 pdb) a été obtenupar amplification PCR sur ADN génomiqueà l’aide des mêmes amorces LAX3F etLAX3R.
2) CgAUX1 et CgLAX3 appartiennentà une petite famille de gènesLe nombre de gènes AUX-LAX chez C.
glauca a été estimé par la technique duSouthern blot. Trois sondes différentes ontété utilisées pour les hybridations : unesonde aspécifique dessinée dans une desrégions les plus conservées des gènesAUX-LAX (exon VII) et deux sondesspécifiques dessinées dans les régionsUTR 3’ de CgAUX1 et CgLAX3respectivement. La sonde aspécifique,utilisée en conditions non-stringentes, aaccroché un nombre limité de bandes quiont toutes pu être identifiées grâce àl’utilisation des sondes spécifiques,utilisées en conditions stringentes (Figure23A). Pour chaque hybridation, auminimum deux répétitions ont été
effectuées. L’absence de bande non-identifiée et le fait que la base de donnéesEST (2700 étiquettes exprimées dans lesracines et les nodules ; Hocher et al, 2006)générée dans notre équipe ne contientaucune nouvelle séquence AUX-LAXsuggèrent que la famille multigénique necontient que ces deux gènes.
3) Structure des gènes CgAUX1 etCgLAX3L’alignement des séquences
génomiques et des ADNc a permis demettre en évidence la présence de 8 exonset 7 introns pour chacun des deux gènesde l’ATG au codon stop. La taille et laposition des exons est très conservéeentre CgLAX3 et CgAUX1 (Figure 23B) ; parcontre la taille des introns n’est pasconservée. Chez A. thaliana, le nombredes introns varie dans la famille AUX-LAX :7 introns pour AtAUX1 et AtLAX1, 5introns pour AtLAX2 et 6 introns pourAtLAX3. Pour la famille AUX-LAX d’A.thaliana, la taille des 5 premiers exons estconservée puis les tailles divergent pourles derniers exons. Les gènes AUX-LAX deMedicago truncatula présentent la mêmestructure que ceux de C. glauca ce quisuggère une origine évolutive commune(Schnabel et Frugoli, 2004). Chez le riz, lenombre d’exons est légèrement différent :3 gènes présentent 7 exons et deux autresgènes en possèdent 5 ou 2. Néanmoins, laposition des introns est très conservéeentre Arabidopsis, M. truncatula, C. glaucaet le riz ce qui suggère que la structure de
Figure 23. La famille AUX-LAX de CasuarinaglaucaLa famille AUX-LAX de C. glauca a été identifiéeen amplifiant de l’ADN génomique à l’aide decouples d’amorces dégénérées. L’hybridation enSouthern blot a été réalisée en utilisant unesonde conservée (1), une sonde spécifique deCgAUX1 (2) ou une sonde spécifique de CgLAX3(3). La sonde conservée n’a pas permis demettre en évidence de nouvelles bandes nonidentifiables comme appartenant à CgAUX1 ouCgLAX3 (A). Structure des gènes CgAUX1 etCgLAX3 par comparaison avec les gènesAtAUX1 et AtLAX3. Exons : boîtes grises ;introns : lignes noires (B).

25
ces gènes est apparue très tôt au cours del’évolution, probablement avant ladivergence des mono- et desdicotylédones.
4) Séquences protéiques déduitesLes phases ouvertes de lecture (ORF)
des ADNc de CgLAX3 et CgAUX1 ont unetaille respective de 1395 pdb et 1440 pdb.Les séquences protéiques déduites ontune taille de 465 acides aminés pour LAX3(ce qui correspond à une protéine de 52,4kDa) et 480 acides aminés pour AUX1(54,0 kDa). L’identité entre les deuxséquences est de 76,2%. Les différencess’observent principalement dans les 40premiers acides aminés (Figure 24A). Lesdeux protéines sont riches en acidesaminés hydrophobes (principalementalanine et leucine qui représentent 20% ennombre des acides aminés). Les acidesaminés dont la mutation entraîne uneperte de fonction de la protéine AtAUX1(Swarup et al, 2004) sont tous conservés
chez CgLAX3 et tous sauf un sontconservés chez CgAUX1 (Figure 24A).
Un arbre phylogénique (Figure 24B) aété réalisé sur la base d’alignement deséquences protéiques. Les séquencesprotéiques déduites CgLAX3 et CgAUX1ont été comparées aux séquencesprotéiques d’un représentant desdifférentes classes de la famille desperméases à auxine et acides aminés(AAAP pour auxin and amino acidpermeases) dont font partie les gènesAUX-LAX. Cette famille contientégalement les perméases à acides aminés(AAP pour amino acid permeases), lestransporteurs de lysine et histidine (LHTpour lysine histidine transporters) et lestransporteurs d’acides aminésaromatiques et d’acides aminés neutres(ANT pour aromatic and neutral aminoacid transporter class). La séquence dutransporteur de saccharose (AtSUC1) a étéutilisée comme séquence de référencepour la construction de l’arbre. L’analyse
Figure 24. Comparaison des séquencesprotéiques déduites de CgAUX1 et CgLAX3Alignement des protéines AtAUX1, CgAUX1 etCgLAX3, les acides aminés importants pour lafonction de transporteur d’influx d’auxine(Swarup et al., 2004) sont indiqués par uneflèche. La flèche noire indique la position del’acide aminé non conservé chez CgAUX1 (A).Arbre phylogénétique contenant les protéinesAUX-LAX de Casuarina glauca et Arabidopsisainsi que les transporteurs d’acides aminésAtANT1, AtAAP1 et AtLHT1. L’arbre a étéenraciné avec le transporteur de saccharoseAtSUC1 (B). Les nombres indiquent le résultat del’analyse Bootstrap (n = 100).
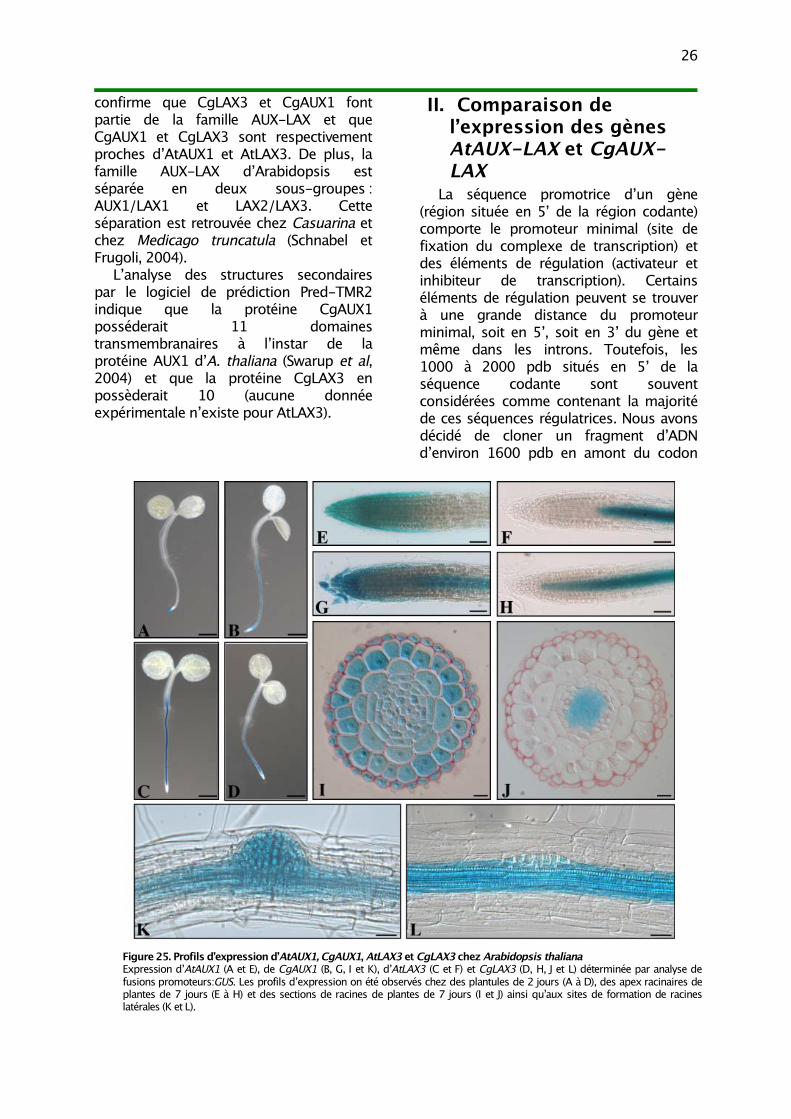
26
confirme que CgLAX3 et CgAUX1 fontpartie de la famille AUX-LAX et queCgAUX1 et CgLAX3 sont respectivementproches d’AtAUX1 et AtLAX3. De plus, lafamille AUX-LAX d’Arabidopsis estséparée en deux sous-groupes :AUX1/LAX1 et LAX2/LAX3. Cetteséparation est retrouvée chez Casuarina etchez Medicago truncatula (Schnabel etFrugoli, 2004).
L’analyse des structures secondairespar le logiciel de prédiction Pred-TMR2indique que la protéine CgAUX1posséderait 11 domainestransmembranaires à l’instar de laprotéine AUX1 d’A. thaliana (Swarup et al,2004) et que la protéine CgLAX3 enpossèderait 10 (aucune donnéeexpérimentale n’existe pour AtLAX3).
II. Comparaison del’expression des gènesAtAUX-LAX et CgAUX-LAX
La séquence promotrice d’un gène(région située en 5’ de la région codante)comporte le promoteur minimal (site defixation du complexe de transcription) etdes éléments de régulation (activateur etinhibiteur de transcription). Certainséléments de régulation peuvent se trouverà une grande distance du promoteurminimal, soit en 5’, soit en 3’ du gène etmême dans les introns. Toutefois, les1000 à 2000 pdb situés en 5’ de laséquence codante sont souventconsidérées comme contenant la majoritéde ces séquences régulatrices. Nous avonsdécidé de cloner un fragment d’ADNd’environ 1600 pdb en amont du codon
Figure 25. Profils d’expression d’AtAUX1, CgAUX1, AtLAX3 et CgLAX3 chez Arabidopsis thalianaExpression d’AtAUX1 (A et E), de CgAUX1 (B, G, I et K), d’AtLAX3 (C et F) et CgLAX3 (D, H, J et L) déterminée par analyse defusions promoteurs:GUS. Les profils d’expression on été observés chez des plantules de 2 jours (A à D), des apex racinaires deplantes de 7 jours (E à H) et des sections de racines de plantes de 7 jours (I et J) ainsi qu’aux sites de formation de racineslatérales (K et L).

27
d’initiation de la traduction (ATG) desgènes CgLAX3 et CgAUX1 respectivement.Cette séquence sera nommée« promoteur » par la suite.
Les séquences situées en amont del’ATG ont été clonées par la technique dela marche sur ADN génomique. Cettetechnique a permis de déterminer laséquence des promoteurs sur unelongueur de 2229 pdb et 1675 pdb enamont de l’ATG pour CgAUX1 et CgLAX3respectivement. Les études de promoteuront par la suite été réalisées en utilisantdes séquences de 1685 pdb et 1650 pdbpour CgAUX1 et CgLAX3 respectivement.
Les promoteurs de CgAUX1 et CgLAX3ont été clonés dans le vecteur pBI101.3 enamont du gène rapporteur GUS (β-glucuronidase) et dans le pBI101.mGFP5ER(Svistoonoff, 2003) en amont du gènerapporteur GFP (green fluorescent protein).Les études d’expression ont tout d’abordété réalisées en système hétérologue chezA. thaliana. Pour cela, les vecteurs binairesont été introduits chez Agrobacteriumtumefaciens afin de transformer desplants d’A. thaliana écotype Col0. Leniveau de fluorescence de la GFP dans lesplantes T1 étant très faible pour CgLAX3et non-détectable pour CgAUX1, le gènerapporteur GUS a été utilisé pour lesétudes d’expression. L’analyse des lignéesT3 homozygotes (environ 5 lignées pourchaque construction) a révélé des profilsd’expression similaires entre les gènesd’Arabidopsis et de C. glauca.
L’expression de CgAUX1 a été détectéedans les apex racinaires, les primordias deracine latérale et le système vasculaire(Figure 25B, G, I et K). AtAUX1 est exprimédans les apex racinaires et les primordiasde racines latérales (Figure 25A et E ;Marchant et al, 2002). Les profilsd’expression de CgAUX1 et AtAUX1 sontrelativement similaires chez A. thaliana.L’expression de CgLAX3 a été détectéeuniquement dans le système vasculaire dela racine (Figure 25D, H, J et L). AtLAX3 estexprimé dans le système vasculaire (Figure25C et F) et dans les cellules du cortex etde l’épiderme situées en face duprimordium de racine latérale (donnée nonmontrée – voir partie III). Là encore, une
grande similarité est observée entre lesprofils d’expression de CgLAX3 et AtLAX3chez A. thaliana.
La relative conservation des profilsd’expression entre CgAUX1 et AtAUX1 etentre CgLAX3 et AtLAX3 tend à montrerque les gènes de C. glauca etd’Arabidopsis sont bien homologues. Celasuggère qu’ils pourraient être impliquésdans les mêmes mécanismesdéveloppementaux chez ces deuxespèces. De plus ces résultats montrentque les mécanismes de régulationtranscriptionnelle sont conservés entre lesBrassicacées et les Casuarinacées. En effet,les Brassicacées appartiennent au cladedes Rosid II et les Casuarinacées au cladedes Rosid I (Figure 26).
III. Analyse fonctionnelleLes gènes CgAUX1 et CgLAX3
présentent un fort pourcentage d’identitéavec les gènes AtAUX1 et AtLAX3respectivement. De plus, l’expression deces gènes en système hétérologue chezArabidopsis montre que leurs profilsd’expression sont très conservés. Cesrésultats suggèrent que les protéinesCgAUX1 et CgLAX3 ont la même fonctionque leurs homologues chez Arabidopsis.Afin de déterminer une éventuelle
Brassicacées
Casuarinacées
Figure 26. Relation phylogénique entre Casuarinacéeset BrassicacéesCasuarina glauca et Arabidopsis thaliana sont des plantesrelativement éloignées phylogénétiquement, appartenant àdeux sous-clades différents.

28
conservation de leur fonction, les gènesCgAUX-LAX de C. glauca ont été introduitschez les mutants aux1 et lax3 d’A.thaliana afin de tenter de complémenterleur phénotype.
1) Complémentation fonctionnelle dumutant aux1Les séquences ADNc de CgAUX1 et
CgLAX3 ont été clonées dans le vecteurpMOG AUX1 ORFL entre les séquencespromotrices (1,7kb) et terminatrices(0,3kb) de AtAUX1. L’objectif était deproduire les protéines CgAUX1 et CgLAX3dans les tissus d’A. thaliana où estnormalement produite la protéine AtAUX1.Ces constructions ont été introduites dansdes plantes mutantes aux1 d’Arabidopsispar transformation génétique. Le mutantaux1 présente normalement unecroissance agravitropique de la racine(Marchant et al, 1999). L’analyse duphénotype a été réalisée sur les plantesT1. Deux témoins ont été réalisés pourcette expérience. Tout d’abord la phasecodante d’AtAUX1 a été placé sousdépendance de son propre promoteur etterminateur afin de constituer un témoinpositif de l’expérience. Ensuite, un vecteurvide ne contenant que le promoteur et leterminateur d’AtAUX1 a été utilisé commetémoin négatif.
L’introduction de la construction necontenant que le promoteur et leterminateur d’AtAUX1 dans le mutantaux1 ne permet pas de restaurer lephénotype sauvage, les plantes présententtoujours un phénotype agravitropique de
la racine (Figure 27). L’expressiond’AtAUX1 dans un fond génétique mutantaux1 permet de restaurer un phénotypesauvage gravitropique de la racine (Figure27). Dans les mêmes conditionsd’expérimentation, l’expression deCgAUX1 sous dépendance du promoteuret du terminateur d’AtAUX1 permet derestaurer un phénotype sauvagegravitropique (Figure 27). À l’inverse,l’expression de CgLAX3 sous dépendancedes séquences régulatrices d’AtAUX1 nerestaure pas de phénotype sauvage auxmutants aux1 (Figure 27).
La réponse gravitropique nécessite unerapide relocalisation des flux d’auxinedans la coiffe et les cellules de l’épiderme(Swarup et al, 2005) et seul untransporteur fonctionnel d’auxine peutassurer cette fonction. La protéineCgAUX1 est donc un transporteurd’auxine de fonction équivalente à celled’AtAUX1. En revanche, l’expression deCgLAX3 ne restaure pas le phénotypesauvage des mutants aux1 ce qui peutsuggérer que : i) la construction contientune erreur qui peut empêcher laproduction de l’ARNm ou ii) la protéineCgLAX3 n’est pas fonctionnellementéquivalente à AtAUX1. Le séquençage desconstructions au niveau des jonctionspromoteur-séquence codante et séquencecodante-terminateur n’a mis en évidenceaucune erreur. De plus, des analyses deRT-PCR démontrent l’accumulation detranscrits correspondants à CgLAX3 ce quiindique que la construction ne contientpas d’erreur empêchant la productiond’ARNm (Figure 28). Il semble donc que laprotéine CgLAX3 n’est pasfonctionnellement équivalente à AtAUX1.
Figure 27. Phénotype gravitropique des mutants aux1-22 complémentés avec les gènes de Casuarina glaucaLe phénotype gravitropique de n plantes T1 a étédéterminé en exerçant un stimulus gravitropique (rotationdes boîtes à 90°). Les plantes ont été groupées en 8classes en fonction de la réorientation des apex racinaires24 heures après stimulus. Le pourcentage de plantes danschaque groupe est indiqué par une barre orientée. Figure 28. Détection des ARNm de CgLAX3 chez les
mutants aux1-22 complémentés par CgLAX3Les ARNm ont été détectés par RT-PCR chez des plantessauvages (Col0) et trois lignées (L1 à L3) de mutants aux1contenant la construction ProAtAUX1:CgLAX3. L’amplificationdes cDNA (+) et les témoins ARN (-) sont présentés ainsique les témoins (vecteur utilisé pour la transformation – C)ou eau. Le gène codant l’actine (AtACT2) a été utilisécomme témoin positif.
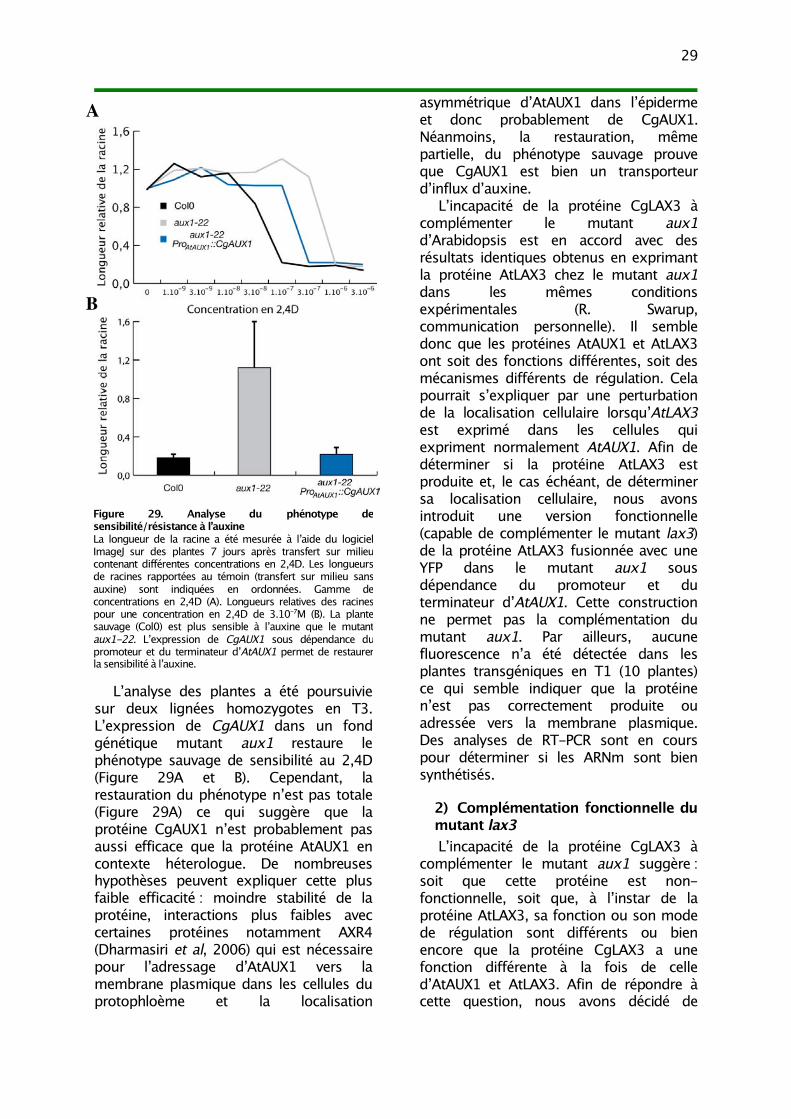
29
L’analyse des plantes a été poursuiviesur deux lignées homozygotes en T3.L’expression de CgAUX1 dans un fondgénétique mutant aux1 restaure lephénotype sauvage de sensibilité au 2,4D(Figure 29A et B). Cependant, larestauration du phénotype n’est pas totale(Figure 29A) ce qui suggère que laprotéine CgAUX1 n’est probablement pasaussi efficace que la protéine AtAUX1 encontexte héterologue. De nombreuseshypothèses peuvent expliquer cette plusfaible efficacité : moindre stabilité de laprotéine, interactions plus faibles aveccertaines protéines notamment AXR4(Dharmasiri et al, 2006) qui est nécessairepour l’adressage d’AtAUX1 vers lamembrane plasmique dans les cellules duprotophloème et la localisation
asymmétrique d’AtAUX1 dans l’épidermeet donc probablement de CgAUX1.Néanmoins, la restauration, mêmepartielle, du phénotype sauvage prouveque CgAUX1 est bien un transporteurd’influx d’auxine.
L’incapacité de la protéine CgLAX3 àcomplémenter le mutant aux1d’Arabidopsis est en accord avec desrésultats identiques obtenus en exprimantla protéine AtLAX3 chez le mutant aux1dans les mêmes conditionsexpérimentales (R. Swarup,communication personnelle). Il sembledonc que les protéines AtAUX1 et AtLAX3ont soit des fonctions différentes, soit desmécanismes différents de régulation. Celapourrait s’expliquer par une perturbationde la localisation cellulaire lorsqu’AtLAX3est exprimé dans les cellules quiexpriment normalement AtAUX1. Afin dedéterminer si la protéine AtLAX3 estproduite et, le cas échéant, de déterminersa localisation cellulaire, nous avonsintroduit une version fonctionnelle(capable de complémenter le mutant lax3)de la protéine AtLAX3 fusionnée avec uneYFP dans le mutant aux1 sousdépendance du promoteur et duterminateur d’AtAUX1. Cette constructionne permet pas la complémentation dumutant aux1. Par ailleurs, aucunefluorescence n’a été détectée dans lesplantes transgéniques en T1 (10 plantes)ce qui semble indiquer que la protéinen’est pas correctement produite ouadressée vers la membrane plasmique.Des analyses de RT-PCR sont en courspour déterminer si les ARNm sont biensynthétisés.
2) Complémentation fonctionnelle dumutant lax3L’incapacité de la protéine CgLAX3 à
complémenter le mutant aux1 suggère :soit que cette protéine est non-fonctionnelle, soit que, à l’instar de laprotéine AtLAX3, sa fonction ou son modede régulation sont différents ou bienencore que la protéine CgLAX3 a unefonction différente à la fois de celled’AtAUX1 et AtLAX3. Afin de répondre àcette question, nous avons décidé de
A
B
Figure 29. Analyse du phénotype desensibilité/résistance à l’auxineLa longueur de la racine a été mesurée à l’aide du logicielImageJ sur des plantes 7 jours après transfert sur milieucontenant différentes concentrations en 2,4D. Les longueursde racines rapportées au témoin (transfert sur milieu sansauxine) sont indiquées en ordonnées. Gamme deconcentrations en 2,4D (A). Longueurs relatives des racinespour une concentration en 2,4D de 3.10-7M (B). La plantesauvage (Col0) est plus sensible à l’auxine que le mutantaux1-22. L’expression de CgAUX1 sous dépendance dupromoteur et du terminateur d’AtAUX1 permet de restaurerla sensibilité à l’auxine.
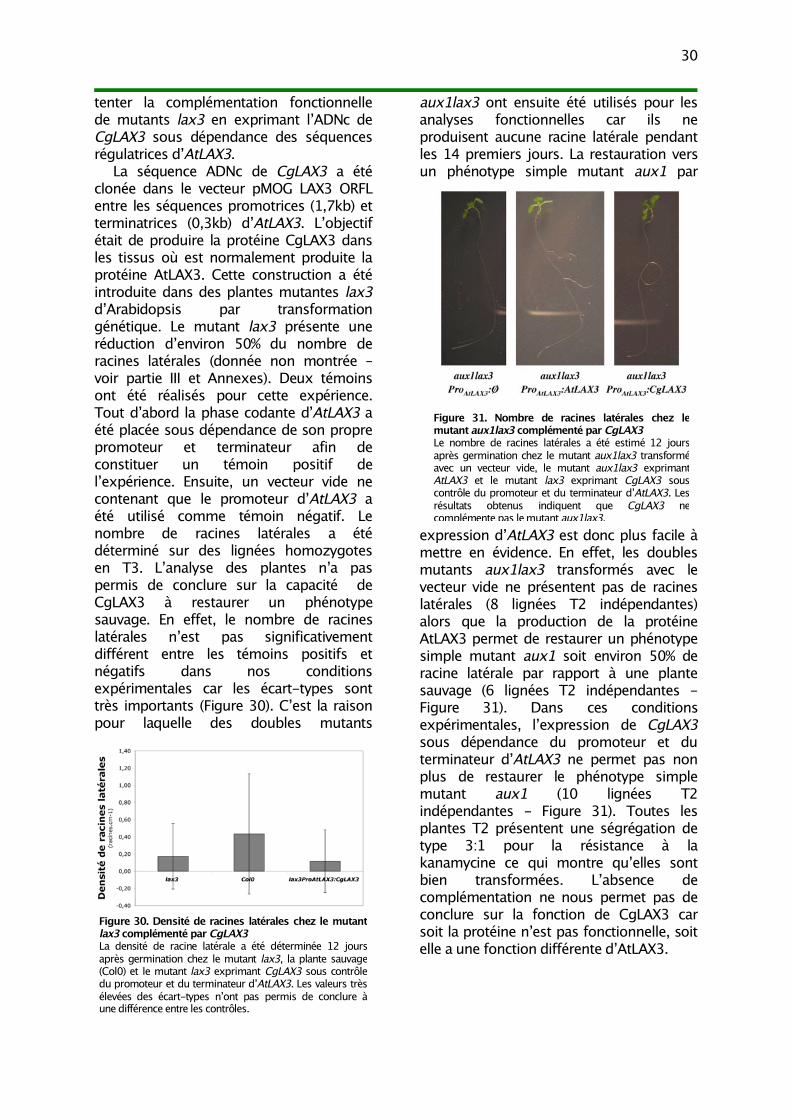
30
tenter la complémentation fonctionnellede mutants lax3 en exprimant l’ADNc deCgLAX3 sous dépendance des séquencesrégulatrices d’AtLAX3.
La séquence ADNc de CgLAX3 a étéclonée dans le vecteur pMOG LAX3 ORFLentre les séquences promotrices (1,7kb) etterminatrices (0,3kb) d’AtLAX3. L’objectifétait de produire la protéine CgLAX3 dansles tissus où est normalement produite laprotéine AtLAX3. Cette construction a étéintroduite dans des plantes mutantes lax3d’Arabidopsis par transformationgénétique. Le mutant lax3 présente uneréduction d’environ 50% du nombre deracines latérales (donnée non montrée –voir partie III et Annexes). Deux témoinsont été réalisés pour cette expérience.Tout d’abord la phase codante d’AtLAX3 aété placée sous dépendance de son proprepromoteur et terminateur afin deconstituer un témoin positif del’expérience. Ensuite, un vecteur vide necontenant que le promoteur d’AtLAX3 aété utilisé comme témoin négatif. Lenombre de racines latérales a étédéterminé sur des lignées homozygotesen T3. L’analyse des plantes n’a paspermis de conclure sur la capacité deCgLAX3 à restaurer un phénotypesauvage. En effet, le nombre de racineslatérales n’est pas significativementdifférent entre les témoins positifs etnégatifs dans nos conditionsexpérimentales car les écart-types sonttrès importants (Figure 30). C’est la raisonpour laquelle des doubles mutants
aux1lax3 ont ensuite été utilisés pour lesanalyses fonctionnelles car ils neproduisent aucune racine latérale pendantles 14 premiers jours. La restauration versun phénotype simple mutant aux1 par
expression d’AtLAX3 est donc plus facile àmettre en évidence. En effet, les doublesmutants aux1lax3 transformés avec levecteur vide ne présentent pas de racineslatérales (8 lignées T2 indépendantes)alors que la production de la protéineAtLAX3 permet de restaurer un phénotypesimple mutant aux1 soit environ 50% deracine latérale par rapport à une plantesauvage (6 lignées T2 indépendantes -Figure 31). Dans ces conditionsexpérimentales, l’expression de CgLAX3sous dépendance du promoteur et duterminateur d’AtLAX3 ne permet pas nonplus de restaurer le phénotype simplemutant aux1 (10 lignées T2indépendantes - Figure 31). Toutes lesplantes T2 présentent une ségrégation detype 3:1 pour la résistance à lakanamycine ce qui montre qu’elles sontbien transformées. L’absence decomplémentation ne nous permet pas deconclure sur la fonction de CgLAX3 carsoit la protéine n’est pas fonctionnelle, soitelle a une fonction différente d’AtLAX3.
Figure 30. Densité de racines latérales chez le mutantlax3 complémenté par CgLAX3La densité de racine latérale a été déterminée 12 joursaprès germination chez le mutant lax3, la plante sauvage(Col0) et le mutant lax3 exprimant CgLAX3 sous contrôledu promoteur et du terminateur d’AtLAX3. Les valeurs trèsélevées des écart-types n’ont pas permis de conclure àune différence entre les contrôles.
Figure 31. Nombre de racines latérales chez lemutant aux1lax3 complémenté par CgLAX3Le nombre de racines latérales a été estimé 12 joursaprès germination chez le mutant aux1lax3 transforméavec un vecteur vide, le mutant aux1lax3 exprimantAtLAX3 et le mutant lax3 exprimant CgLAX3 souscontrôle du promoteur et du terminateur d’AtLAX3. Lesrésultats obtenus indiquent que CgLAX3 necomplémente pas le mutant aux1lax3.

31
IV. Discussion
1) Une petite famille multigéniqueAUX-LAX chez C. glaucaNous avons isolé des homologues des
gènes AUX-LAX chez C. glauca.Différentes approches ont été menéespour tenter de déterminer le nombre degènes de la famille AUX-LAX de C. glauca.Tout d’abord, une approche paramplification PCR sur ADN génomique àl’aide d’amorces dégénérées dessinéesdans des régions très conservées nous apermis d’identifier deux gènes. Lesséquences correspondant aux amorces setrouvent dans les exons 3 (AD1 et AF2), 4(AD2), 5 (AF3) et 6 (AD3 et AD4) etcouvrent donc une grande partie desséquences conservées du gène. Ensuite,une sonde correspondant à une séquencetrès conservée de la famille (située dans laquasi-totalité de l’exon 7) a été utiliséepour l’hybridation en Southern blot. Pasplus de deux bandes n’ont été obtenuespour chaque digestion de l’ADN. Toutesces expériences ainsi que l’absence denouvelles séquences AUX-LAX dans lesbanques EST de C. glauca suggèrent quela famille multigénique de C. glauca necontient que deux membres.
La famille AUX-LAX de C. glauca necomporte donc que deux gènes comparéeà celle d’Arabidopsis (4 gènes ; Parry et al,2001b), de Medicago truncatula (5 gènes ;Schnabel et Frugoli, 2004) ou encore duriz (Oryza sativa ssp japonica cv.Nipponbare - 5 gènes ; Tyagi et al, 2004).Néanmoins, le génome de C. glauca estrelativement petit (estimé à environ 290Mpb ; Schwencke et al, 1998) comparé auriz (430 Mpb ; Tyagi et al, 2004) ou à M.truncatula (470 Mpb ; Cannon et al, 2006)mais plus grand que celui d’Arabidopsis(125 Mbp ; Arabidopsis Genome Initiative,2000). Chez Arabidopsis, les gènes AUX-LAX ne sont pas situés dans les régionsdupliquées du génôme ce qui indiquequ’ils proviennent de duplicationsspécifiques de gènes plutôt que de largesduplications chromosomiques.
Les deux homologues des gènes AUX-LAX ont été nommés CgAUX1 et CgLAX3par homologie avec les gènes AtAUX1 et
AtLAX3 respectivement. Des ADNccorrespondant aux deux gènes etcontenant la séquence codante complèteont été obtenus par RACE PCR. La trèsforte identité de séquence (au niveaunucléique et au niveau protéique) de cesgènes avec les gènes connus chez d’autresespèces suggère qu’ils appartiennenteffectivement à la famille destransporteurs d’influx d’auxine. Parailleurs, la structure des gènes CgLAX3 etCgAUX1 est identique à celle du gèneAtAUX1 (taille et position des exons) cequi indique qu’il existe une origineévolutive commune de ces séquences. Deplus, les acides aminés dont la mutationentraîne une perte de fonction de laprotéine AtAUX1 chez A. thaliana (Swarupet al, 2004) sont conservés à l’exceptiond’un dans la séquence de CgAUX1.
2) Divergence fonctionnelle dans lafamille AUX-LAXLes analyses phylogénétiques des
protéines AUX-LAX suggèrent qu’il existedeux groupes fonctionnels distincts chezArabidopsis ainsi que chez M. truncatula(Schnabel et Frugoli, 2004). Les deuxprotéines identifiées chez C. glaucaappartiennent à chacun de ces deuxgroupes potentiellement distincts. Cettedifférence fonctionnelle est égalementobservée lors des expériences decomplémentation des mutants aux1 parles protéines CgAUX1 et CgLAX3/AtLAX3.En effet, les protéines AtLAX3/CgLAX3sont incapables de restaurer le phénotypede ce mutant. Pourtant, la protéine AtLAX3est un transporteur d’influx d’auxine àhaute affinité comme AtAUX1 (M. Bennett,communication personnelle). Par ailleurs,ces deux groupes de gènes sont exprimésdans des tissus différents. En effet,AtAUX1 est exprimé dans l’apex racinaireet les primordia de racine latérale alorsqu’AtLAX3 est exprimé dans le systèmevasculaire et les cellules situées en facedes primordia. Cela indique que ces deuxgènes sont impliqués dans desmécanismes développementaux distincts :gravitropisme, initiation et organisation duprimordium de racine latérale pourAtAUX1 et émergence pour AtLAX3. Il est

32
fort probable que la spécialisation desprofils d’expression d’AtAUX1 et AtLAX3s’est associée à la divergence fonctionnellede ces protéines au cours de l’évolution.En tout cas, cette divergence a eu lieuavant la séparation entre les Casuarinacéeset les Brassicacées.
3) Bases moléculaires de ladivergenceLa régulation de ces protéines a
probablement divergé au cours de laspécialisation fonctionnelle. Afin depréciser les mécanismes moléculairesresponsables de ces différences, nousavons essayé de déterminer pourquoi laprotéine AtLAX3 est incapable decomplémenter le mutant aux1. Pour cela,nous avons introduit une fusionfonctionnelle AtLAX3:YFP dans le mutantaux1. L’absence de fluorescence observéesemble indiquer une absence deproduction de la protéine, unedégradation rapide ou une mauvaiselocalisation. La séquence codanted’AtLAX3 a été placée entre les séquencesUTR 5’ et 3’ du gène AtAUX1 ce quipermet de conserver une partie desmécanismes de régulationtranscriptionnelle et traductionnelle.Néanmoins, il n’est pas possible d’écarterune régulation de type post-transcriptionnelle par les microARN (Zhanget al, 2007). Il peut également s’agir d’unproblème d’adressage subcellulaire de laprotéine AtLAX3. Des études récentes ontmontré que la protéine AtAUX1 nécessited’interagir avec la protéine AXR4 pour êtrecorrectement localisée dans la membraneplasmique (Figure 32 ; Dharmasiri et al,2006). La mutation axr4 affectespécifiquement la localisation d’AtAUX1, laprotéine AtLAX3 n’est pas affectée (M.Bennett, communication personnelle). Celaindique qu’AtLAX3 n’intéragitprobablement pas avec AXR4. À ce jour,
aucun partenaire d’AtLAX3 nécessairepour sa localisation sur la membrane n’aété identifié. Cependant, un tel partenairene serait a priori pas exprimé dans lesmêmes tissus qu’AtAUX1. Cela pourraitsuffire à expliquer le mauvais adressaged’AtLAX3 sous dépendance du promoteurd’AtAUX1. L’absence de fluorescenceindique que la protéine ne s’accumule pas
dans le réticulum endoplasmique, ce quipourrait se produire en l’absenced’adressage vers la membrane. Laprotéine AtLAX3 pourrait donc êtredégradée par la cellule lorsque salocalisation n’est pas correcte. L’analyse encours des plantes aux1 ProAtAUX1::AtLAX3-YFP devrait permettre de répondre à cesinterrogations.
Afin de préciser la différencefonctionnelle entre AtAUX1 et AtLAX3, lacomplémentation du mutant lax3 parl’expression de la protéine AtAUX1 a été
initiée. Le clonage d’une versionfonctionnelle de la protéine AUX1:YFPentre les séquences régulatrices(promoteur et terminateur) d’AtLAX3 esten cours. L’introduction de cetteconstruction chez des mutants lax3 (ou
double mutants aux1lax3 pour faciliterl’analyse phénotypique) nous permettra dedéterminer si AtAUX1 peut remplacerAtLAX3 et de déterminer la localisationcellulaire de la protéine.
Figure 32. AXR4 est nécessaire pour permettre lalocalisation membranaire d’AUX1Trois modèles pour le rôle d’AXR4 dans l’adressaged’AUX1 : AXR4 joue le rôle de chaperone pour éviterl’aggrégation des ségments hydrophobes d’AUX1 (1),AXR4 catalyse une modification post-traductionnelled’AUX1 pour permettre l’adressage (2) ou AXR4 modifie lacomposition lipidique de la membrane pour réguler lalocalisation d’AUX1 (3). ER : réticulum endoplasmique.D’après Hobbie, 2006.

33
PARTIE II - ÉTUDE DU RÔLE DU TRANSPORT D’INFLUX AU COURSDU DÉVELOPPEMENT DU NODULE ACTINORHIZIEN
I. IntroductionUn rôle de l’auxine au cours de la
symbiose actinorhizienne a longtemps étésuggéré car la bactérie Frankia produit del’auxine (Wheeler et al, 1984 ; Hammad etal, 2003). De plus, les nodules contiennentdes quantités importantes d’auxine parrapport aux racines non infectées (Wheeleret al, 1979). Nous avons isolé deux gènesde la famille AUX-LAX chez Casuarinaglauca. L’un d’entre eux, nommé CgAUX1,est fonctionnellement équivalent àAtAUX1. Nous nous sommes intéressé aurôle que pouvaient jouer ces transporteursau cours de la symbiose actinorhizienne.Pour cela, nous avons analysé le profild’expression des gènes CgAUX1 etCgLAX3 au cours de la symbiose C.glauca-Frankia et déterminé l’impact del’inhibition du transport d’influx d’auxinesur la nodulation.
II. Analyse de l’expressionde CgAUX1 et CgLAX3
Le profil d’expression de CgAUX1 etCgLAX3 a été analysé par différentestechniques afin de déterminer si ces gènessont exprimés au cours de l’infection parla bactérie Frankia et du développementdu nodule actinorhizien. Les constructionspromoteur-GUS utilisées pour l’analyse del’expression en système hétérologue chezArabidopsis ont été utilisées pourtransformer C. glauca et Allocasuarinaverticillata. Le protocole de transformationgénétique de C. glauca est plus long quecelui d’A. verticillata et n’aboutit pasforcément à l’obtention de plantestransgéniques. Par contre, la nodulationdes plants d’A. verticillata est beaucoupplus longue à obtenir et les étapesprécoces de la nodulation sont difficiles àobserver. Une lignée transgénique
contenant le promoteur de CgAUX1 a étéobtenue chez C. glauca et deux lignéeschez A. verticillata. Quatre lignéestransgéniques contenant le promoteur deCgLAX3 ont été obtenues chez A.verticillata, par contre aucune lignée n’apu être régénérée chez C. glauca. Enparallèle, des analyses du niveaud’expression des gènes ont été réaliséespar RT-PCR quantitative en réponse à destraitements auxiniques et à l’infection.
1) Expression non-symbiotiqueDans un premier temps, l’expression
des gènes CgAUX1 et CgLAX3 a étéanalysée par RT-PCR non-quantitative afinde vérifier dans quels organes ces gènessont exprimés. Les transcrits des deuxgènes ont été détectés dans les partiesaériennes et les racines non-infectées(Figure 33). L’analyse des lignéespromoteur:GUS montre que CgAUX1 est
exprimé dans les apex racinaires, lesprimordia de racine latérale et le systèmevasculaire de la racine et de la feuille(Figure 34). CgLAX3 est expriméuniquement dans le système vasculaire dela racine. Aucune expression de CgLAX3n’est observée en face des primordia deracine latérale (Figure 34). Les profils
Figure 33. Détection des ARNm de CgAUX1 et CgLAX3Les ARNm de CgAUX1 et CgLAX3 ont été détectés dans lesdifférents organes testés : nodules, parties aériennes etracines.
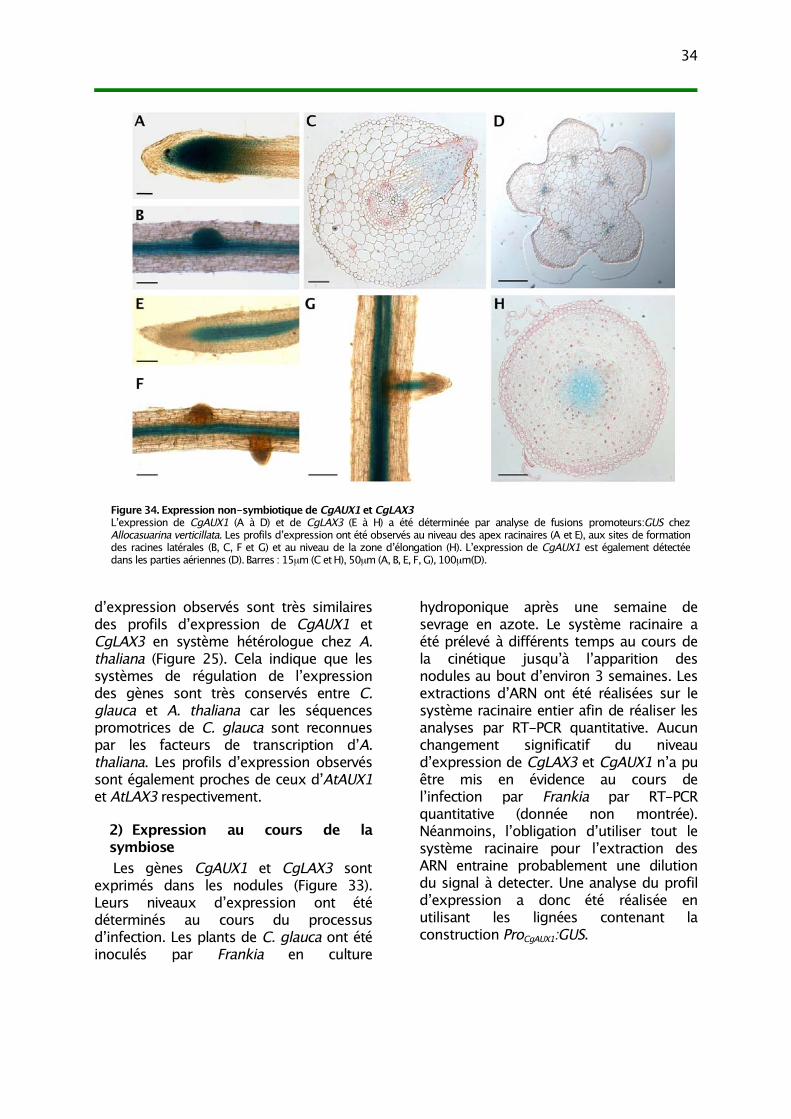
34
d’expression observés sont très similairesdes profils d’expression de CgAUX1 etCgLAX3 en système hétérologue chez A.thaliana (Figure 25). Cela indique que lessystèmes de régulation de l’expressiondes gènes sont très conservés entre C.glauca et A. thaliana car les séquencespromotrices de C. glauca sont reconnuespar les facteurs de transcription d’A.thaliana. Les profils d’expression observéssont également proches de ceux d’AtAUX1et AtLAX3 respectivement.
2) Expression au cours de lasymbioseLes gènes CgAUX1 et CgLAX3 sont
exprimés dans les nodules (Figure 33).Leurs niveaux d’expression ont étédéterminés au cours du processusd’infection. Les plants de C. glauca ont étéinoculés par Frankia en culture
hydroponique après une semaine desevrage en azote. Le système racinaire aété prélevé à différents temps au cours dela cinétique jusqu’à l’apparition desnodules au bout d’environ 3 semaines. Lesextractions d’ARN ont été réalisées sur lesystème racinaire entier afin de réaliser lesanalyses par RT-PCR quantitative. Aucunchangement significatif du niveaud’expression de CgLAX3 et CgAUX1 n’a puêtre mis en évidence au cours del’infection par Frankia par RT-PCRquantitative (donnée non montrée).Néanmoins, l’obligation d’utiliser tout lesystème racinaire pour l’extraction desARN entraine probablement une dilutiondu signal à detecter. Une analyse du profild’expression a donc été réalisée enutilisant les lignées contenant laconstruction ProCgAUX1:GUS.
Figure 34. Expression non-symbiotique de CgAUX1 et CgLAX3L’expression de CgAUX1 (A à D) et de CgLAX3 (E à H) a été déterminée par analyse de fusions promoteurs:GUS chezAllocasuarina verticillata. Les profils d’expression ont été observés au niveau des apex racinaires (A et E), aux sites de formationdes racines latérales (B, C, F et G) et au niveau de la zone d’élongation (H). L’expression de CgAUX1 est également détectéedans les parties aériennes (D). Barres : 15µm (C et H), 50µm (A, B, E, F, G), 100µm(D).
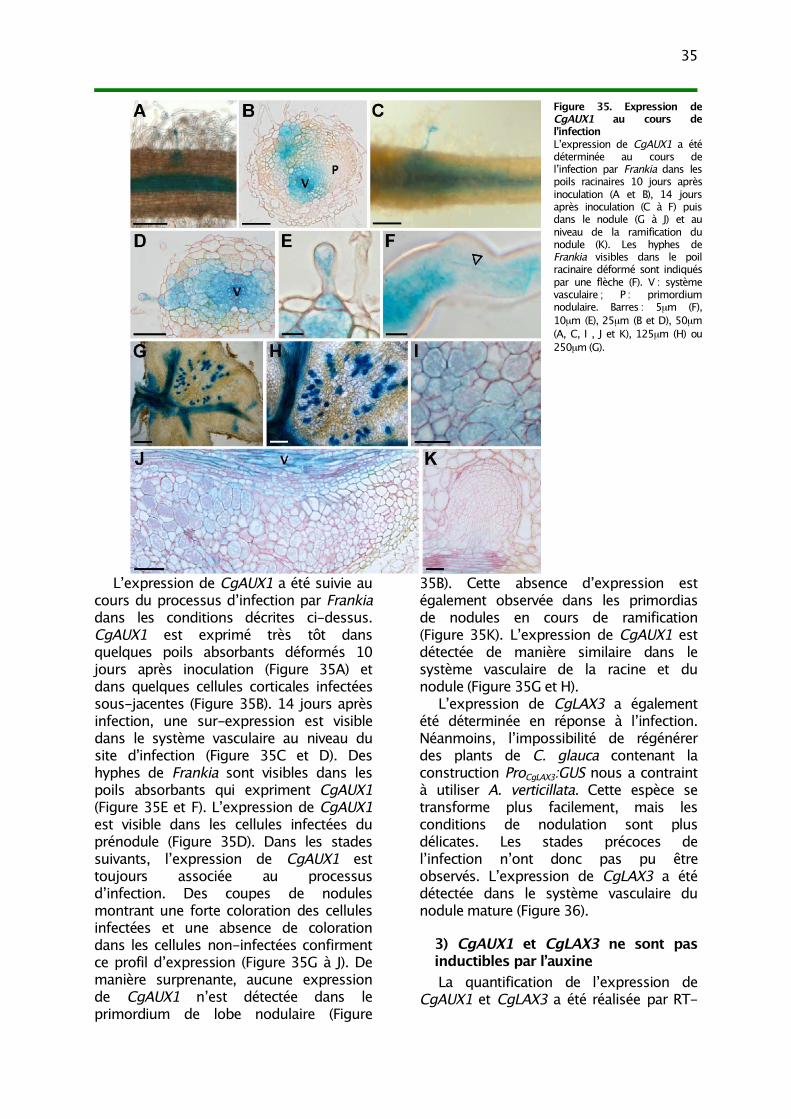
35
L’expression de CgAUX1 a été suivie aucours du processus d’infection par Frankiadans les conditions décrites ci-dessus.CgAUX1 est exprimé très tôt dansquelques poils absorbants déformés 10jours après inoculation (Figure 35A) etdans quelques cellules corticales infectéessous-jacentes (Figure 35B). 14 jours aprèsinfection, une sur-expression est visibledans le système vasculaire au niveau dusite d’infection (Figure 35C et D). Deshyphes de Frankia sont visibles dans lespoils absorbants qui expriment CgAUX1(Figure 35E et F). L’expression de CgAUX1est visible dans les cellules infectées duprénodule (Figure 35D). Dans les stadessuivants, l’expression de CgAUX1 esttoujours associée au processusd’infection. Des coupes de nodulesmontrant une forte coloration des cellulesinfectées et une absence de colorationdans les cellules non-infectées confirmentce profil d’expression (Figure 35G à J). Demanière surprenante, aucune expressionde CgAUX1 n’est détectée dans leprimordium de lobe nodulaire (Figure
35B). Cette absence d’expression estégalement observée dans les primordiasde nodules en cours de ramification(Figure 35K). L’expression de CgAUX1 estdétectée de manière similaire dans lesystème vasculaire de la racine et dunodule (Figure 35G et H).
L’expression de CgLAX3 a égalementété déterminée en réponse à l’infection.Néanmoins, l’impossibilité de régénérerdes plants de C. glauca contenant laconstruction ProCgLAX3:GUS nous a contraintà utiliser A. verticillata. Cette espèce setransforme plus facilement, mais lesconditions de nodulation sont plusdélicates. Les stades précoces del’infection n’ont donc pas pu êtreobservés. L’expression de CgLAX3 a étédétectée dans le système vasculaire dunodule mature (Figure 36).
3) CgAUX1 et CgLAX3 ne sont pasinductibles par l’auxineLa quantification de l’expression de
CgAUX1 et CgLAX3 a été réalisée par RT-
Figure 35. Expression deCgAUX1 au cours del’infectionL’expression de CgAUX1 a étédéterminée au cours del’infection par Frankia dans lespoils racinaires 10 jours aprèsinoculation (A et B), 14 joursaprès inoculation (C à F) puisdans le nodule (G à J) et auniveau de la ramification dunodule (K). Les hyphes deFrankia visibles dans le poilracinaire déformé sont indiquéspar une flèche (F). V : systèmevasculaire ; P : primordiumnodulaire. Barres : 5µm (F),10µm (E), 25µm (B et D), 50µm(A, C, I , J et K), 125µm (H) ou250µm (G).

36
PCR quantitative en réponse à différentstraitements auxiniques. Le gène CgAUX1n’est pas induit par les auxines utilisées(IAA, NAA et PAA) à 1 µM (Figure 37) et 10µM (données non montrées) et le gèneCgLAX3 montre une sous-expression enréponse à certains traitements. Cetteabsence d’induction est observée quel quesoit le statut minéral de la plante (10mMde nitrate, carence en azote d’une semaineet carence en phosphate d’une semaine).Des traitements avec 1 ou 10 µM de NAApendant 16 heures n’ont pas modifié leprofil d’expression déterminé par laconstruction ProCgAUX1 :GUS (donnée nonmontrée). L’utilisation de surnageant deFrankia (dilué au 1/4) n’a pas non plusmodifié le profil d’expression de CgAUX1.Ces différents traitements ont été réalisésen condition de carence en azote (unesemaine). En conclusion, l’auxine n’est pas
un signal suffisant pour induirel’expression de CgAUX1 au cours del’infection et le signal inducteur deCgAUX1 reste à déterminer.
III. L’inhibition du transportd’influx d’auxineperturbe le processus denodulation
Le profil d’expression de CgAUX1suggère que le transport d’influx d’auxineest impliqué dans le processus d’infectionau cours de la mise en place du noduleactinorhizien (Figure 35). Afin de testercette hypothèse, nous avons utilisé l’acide1-naphtoxy acétique (1-NOA) qui est uninhibiteur du transport d’influx d’auxine(Parry et al, 2001a) qui agit parcompétition avec l’auxine (Yang et al,2006). Dans un premier temps, nousavons déterminé l’impact du 1-NOA sur laprotéine CgAUX1 puis l’effet de cettemolécule a été étudié au cours del’infection.
Afin de déterminer si la protéineCgAUX1 est sensible au 1-NOA, nousavons utilisé les mutants aux1-22complémentés par l’expression deCgAUX1 sous dépendance du promoteuret du terminateur d’AtAUX1. Le traitementdes plantes sauvages avec 25 µM de 1-NOA pendant 24 heures avant le testgravitropique entraine une perte duphénotype gravitropique. Les plantesmutantes sont déjà agravitropiques et nesont donc pas affectées par ce traitement(Figure 38A). L’effet du 1-NOA sur lesmutants aux1-22 complémentés parl’expression de CgAUX1 est identique àcelui observé chez les plantes sauvages(Figure 38A). La perte du phénotypegravitropique signifie donc que letransporteur d’influx d’auxine CgAUX1 estsensible au 1-NOA.
L’effet du 1-NOA sur la nodulation aété déterminé en ajoutant 25 µM de 1-NOA au moment de l’inoculation parFrankia puis en suivant l’apparition desprénodules et nodules à l’œil nu tous lesjours jusqu’à ce que toutes les plantessoient nodulées (trois répétitionsbiologiques). Le traitement a entrainé un
Figure 36. Expression de CgLAX3 dans le noduleactinorhizien chez Allocasuarina verticillataL’expression de CgLAX3 est détectée dans le jeunesystème vasculaire du nodule. Barre : 100µm.
0,00
0,20
0,40
0,60
0,80
1,00
1,20
1,40
1,60
1,80
2,00
+N+P -N -P +N+P -N -P
CgAUX1 CgLAX3
Niv
eau
d'e
xp
ress
ion
rela
tif
NT+IAA+NAA+PAA
nd
nd
Figure 37. Induction de CgAUX1 et CgLAX3 par lesauxinesDétermination du niveau d’expression de CgAUX1 etCgLAX3 dans le système racinaire par RT-PCR quantitativeen réponse à des traitements de 1µM en auxines (IAA, NAAou PAA) pendant 24 heures. Les analyses ont été réaliséesen hydroponie sur milieu BD contenant de l’azote et duphosphate (+N+P) ou après sevrage d’une semaine enazote (-N) ou en phosphate (-P). L’expression des gènesest exprimée par rapport aux plantes non-traitées parl’auxine après normalisation par le gène de l’ubiquitine(CgUBI). nd : non déterminé.
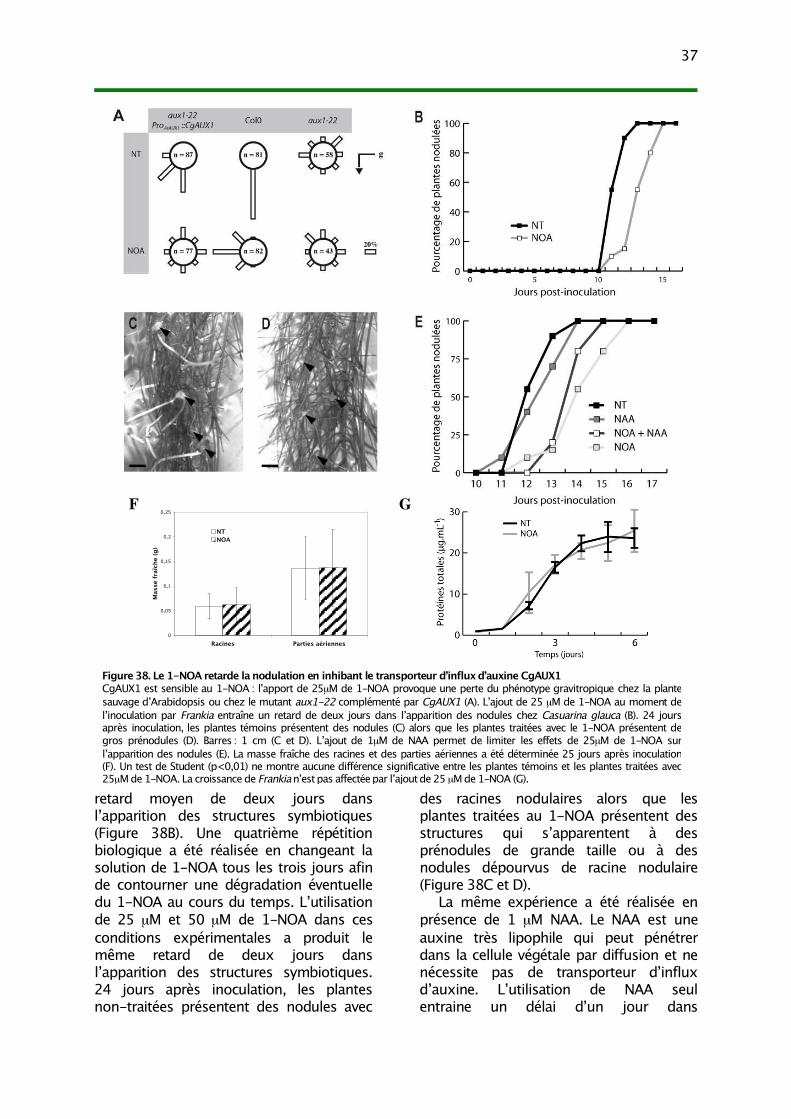
37
retard moyen de deux jours dansl’apparition des structures symbiotiques(Figure 38B). Une quatrième répétitionbiologique a été réalisée en changeant lasolution de 1-NOA tous les trois jours afinde contourner une dégradation éventuelledu 1-NOA au cours du temps. L’utilisationde 25 µM et 50 µM de 1-NOA dans cesconditions expérimentales a produit lemême retard de deux jours dansl’apparition des structures symbiotiques.24 jours après inoculation, les plantesnon-traitées présentent des nodules avec
des racines nodulaires alors que lesplantes traitées au 1-NOA présentent desstructures qui s’apparentent à desprénodules de grande taille ou à desnodules dépourvus de racine nodulaire(Figure 38C et D).
La même expérience a été réalisée enprésence de 1 µM NAA. Le NAA est uneauxine très lipophile qui peut pénétrerdans la cellule végétale par diffusion et nenécessite pas de transporteur d’influxd’auxine. L’utilisation de NAA seulentraine un délai d’un jour dans
Figure 38. Le 1-NOA retarde la nodulation en inhibant le transporteur d’influx d’auxine CgAUX1CgAUX1 est sensible au 1-NOA : l’apport de 25µM de 1-NOA provoque une perte du phénotype gravitropique chez la plantesauvage d’Arabidopsis ou chez le mutant aux1-22 complémenté par CgAUX1 (A). L’ajout de 25 µM de 1-NOA au moment del’inoculation par Frankia entraîne un retard de deux jours dans l’apparition des nodules chez Casuarina glauca (B). 24 joursaprès inoculation, les plantes témoins présentent des nodules (C) alors que les plantes traitées avec le 1-NOA présentent degros prénodules (D). Barres : 1 cm (C et D). L’ajout de 1µM de NAA permet de limiter les effets de 25µM de 1-NOA surl’apparition des nodules (E). La masse fraîche des racines et des parties aériennes a été déterminée 25 jours après inoculation(F). Un test de Student (p<0,01) ne montre aucune différence significative entre les plantes témoins et les plantes traitées avec25µM de 1-NOA. La croissance de Frankia n’est pas affectée par l’ajout de 25 µM de 1-NOA (G).
G
0
0,05
0,1
0,15
0,2
0,25
Racines Parties aériennes
Mas
se f
raîc
he (g
)
NTNOA
F
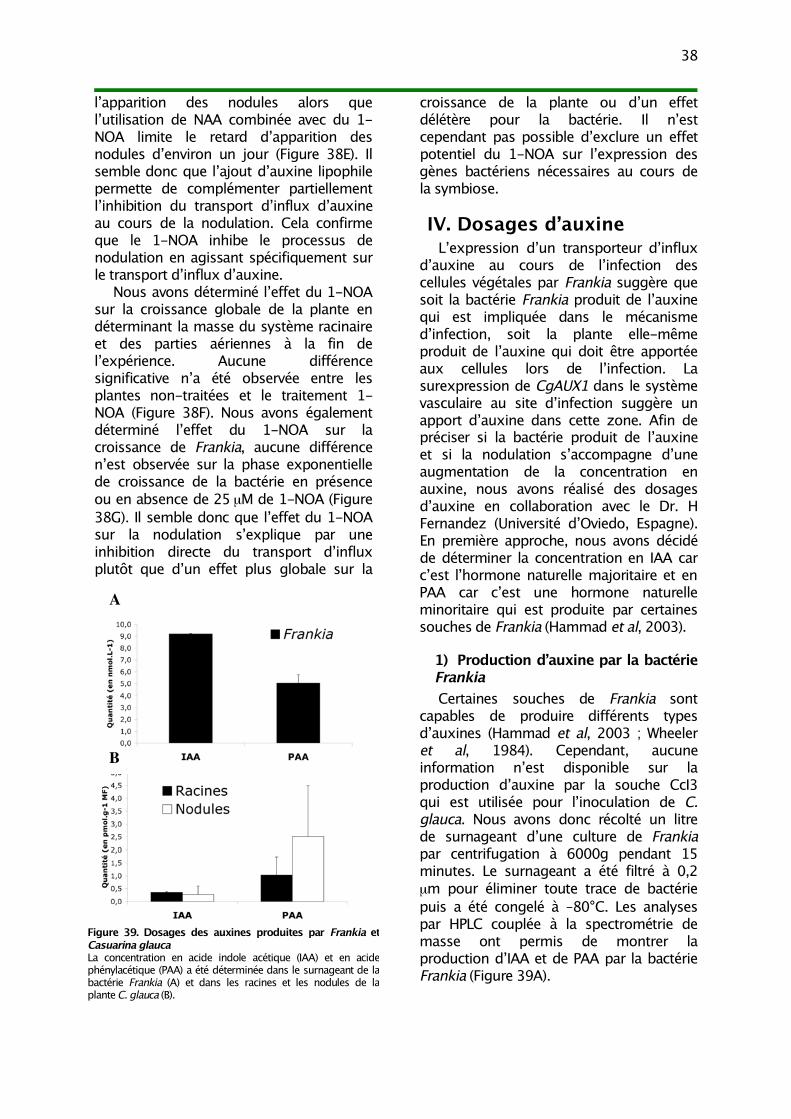
38
l’apparition des nodules alors quel’utilisation de NAA combinée avec du 1-NOA limite le retard d’apparition desnodules d’environ un jour (Figure 38E). Ilsemble donc que l’ajout d’auxine lipophilepermette de complémenter partiellementl’inhibition du transport d’influx d’auxineau cours de la nodulation. Cela confirmeque le 1-NOA inhibe le processus denodulation en agissant spécifiquement surle transport d’influx d’auxine.
Nous avons déterminé l’effet du 1-NOAsur la croissance globale de la plante endéterminant la masse du système racinaireet des parties aériennes à la fin del’expérience. Aucune différencesignificative n’a été observée entre lesplantes non-traitées et le traitement 1-NOA (Figure 38F). Nous avons égalementdéterminé l’effet du 1-NOA sur lacroissance de Frankia, aucune différencen’est observée sur la phase exponentiellede croissance de la bactérie en présenceou en absence de 25 µM de 1-NOA (Figure38G). Il semble donc que l’effet du 1-NOAsur la nodulation s’explique par uneinhibition directe du transport d’influxplutôt que d’un effet plus globale sur la
croissance de la plante ou d’un effetdélétère pour la bactérie. Il n’estcependant pas possible d’exclure un effetpotentiel du 1-NOA sur l’expression desgènes bactériens nécessaires au cours dela symbiose.
IV. Dosages d’auxineL’expression d’un transporteur d’influx
d’auxine au cours de l’infection descellules végétales par Frankia suggère quesoit la bactérie Frankia produit de l’auxinequi est impliquée dans le mécanismed’infection, soit la plante elle-mêmeproduit de l’auxine qui doit être apportéeaux cellules lors de l’infection. Lasurexpression de CgAUX1 dans le systèmevasculaire au site d’infection suggère unapport d’auxine dans cette zone. Afin depréciser si la bactérie produit de l’auxineet si la nodulation s’accompagne d’uneaugmentation de la concentration enauxine, nous avons réalisé des dosagesd’auxine en collaboration avec le Dr. HFernandez (Université d’Oviedo, Espagne).En première approche, nous avons décidéde déterminer la concentration en IAA carc’est l’hormone naturelle majoritaire et enPAA car c’est une hormone naturelleminoritaire qui est produite par certainessouches de Frankia (Hammad et al, 2003).
1) Production d’auxine par la bactérieFrankiaCertaines souches de Frankia sont
capables de produire différents typesd’auxines (Hammad et al, 2003 ; Wheeleret al, 1984). Cependant, aucuneinformation n’est disponible sur laproduction d’auxine par la souche CcI3qui est utilisée pour l’inoculation de C.glauca. Nous avons donc récolté un litrede surnageant d’une culture de Frankiapar centrifugation à 6000g pendant 15minutes. Le surnageant a été filtré à 0,2µm pour éliminer toute trace de bactériepuis a été congelé à –80°C. Les analysespar HPLC couplée à la spectrométrie demasse ont permis de montrer laproduction d’IAA et de PAA par la bactérieFrankia (Figure 39A).
B
A
Figure 39. Dosages des auxines produites par Frankia etCasuarina glaucaLa concentration en acide indole acétique (IAA) et en acidephénylacétique (PAA) a été déterminée dans le surnageant de labactérie Frankia (A) et dans les racines et les nodules de laplante C. glauca (B).

39
2) Accumulation d’auxine au coursde la nodulationLa nodulation du système racinaire de
certaines plantes actinorhiziennes commeAlnus glutinosa, A. serrulata et Myricacerifera est associée avec uneaugmentation de la teneur en auxines(Wheeler et al, 1979). Cependant, il n’ajamais été prouvé si cette auxine estd’origine végétale ou bactérienne. De plus,aucune information n’est connue dans lecas de la symbiose entre C. glauca etFrankia. Nous avons donc récolté lesystème racinaire de plants de C. glauca(conditions non sevrées en azote) et desnodules âgés de deux à trois semaines.Les échantillons ont été placés dansl’azote et envoyés pour des analyses parHPLC couplée à la spectrométrie demasse. Les résultats des dosagesmontrent une accumulation similaire d’IAAdans les deux organes. Le PAA est présenten plus grande quantité que l’IAA dans lesracines et semble être accumulé dans lesnodules (Figure 39B). Néanmoins, denouveaux dosages devront être réalisésafin de confirmer ces résultatspréliminaires.
V. Discussion
1) L’influx d’auxine est associé aumécanisme d’infection par FrankiaLes résultats obtenus suggèrent que
l’influx d’auxine est impliqué dansl’interaction symbiotique entre C. glauca etFrankia. En effet, l’expression de CgAUX1est toujours observée dans les cellulesinfectées par la bactérie symbiotique. Ceprofil d’expression nécessite qu’un signalde Frankia produit au cours de l’infectionentraine une réponse spécifique de laplante. A priori, il ne s’agit pas de l’auxineelle-même car l’application d’auxine ou desurnageant de Frankia n’a pas d’effet surl’expression de CgAUX1.
La production d’un transport d’influxd’auxine suggère que la cellule infectéeprélève de l’auxine dans sonenvironnement immédiat. Plusieurshypothèses sont possibles pour expliquerl’origine de l’auxine perçue par la celluleinfectée. Tout d’abord, l’auxine pourrait
provenir des tissus végétaux adjacents.Dans ce cas, l’expression de CgAUX1permettrait de recruter l’auxine présentedans les tissus végétaux afin de permettrela mise en place du programmesymbiotique. Une autre hypothèse est quela plante prend en charge l’auxineproduite par la bactérie. L’utilisation d’uneprotéine CgAUX1 fusionnée avec uneprotéine fluorescente (GFP ou YFP)permettrait de déterminer la localisationcellulaire de CgAUX1 dans les cellulesinfectées. La détection de la protéine dansla membrane plasmique en périphérie descellules ou dans la membrane plasmiquequi entoure les bactéries permettrait depréciser quelle hypothèse est la plusvraisemblable.
Des dosages ont permis de démontrerque la souche CcI3 de Frankia produit del’auxine. Néanmoins, nous n’avons pasdéterminé si la bactérie produit desauxines au cours de l’infection. Le récentséquençage du génome de Frankia, etnotamment de la souche CcI3, a permisd’identifier les gènes potentiellementimpliqués dans la voie de biosynthèse del’auxine (IAA et PAA ; Normand et al,2006). Le suivi de l’expression de cesgènes au cours de l’infection parhybridation in situ permettrait de mettreen évidence la production d’auxine parFrankia au cours de la symbiose. D’autressouches de Frankia ainsi que denombreuses bactéries de la rhizosphèreont la capacité de produire de l’auxine(Wheeler et al, 1984 ; Lambrecht et al,2000). Cependant, le rôle de l’auxinebactérienne dans les relations plantes-microorganismes n’a jamais été clairementétabli. La production d’auxine par denombreuses rhizobactéries montre quel’auxine seule n’est pas un signalsuffisamment spécifique et doitnécessairement être accompagné d’unsignal spécifique comme les facteurs Nodchez Rhizobium. Dans ce cas, l’auxinepourrait alors agir en synergie avec unsignal symbiotique plus spécifique. Eneffet, la surproduction d’auxine par unesouche mutante de Bradyrhizobiumjaponicum correspond à uneaugmentation de l’efficacité de la

40
nodulation chez les Légumineuses(Kaneshiro et Kwolek, 1985).
Il a été montré qu’AtAUX1 et AtLAX3sont des transporteurs d’auxine à hauteaffinité (Yang et al, 2006; M. Bennett,communication personnelle) cependantleur niveau de spécificité n’est pas connu.Il se pourrait donc qu’un transporteurd’auxine prenne en charge des composésindoliques produits par la bactérie Frankia(Wheeler et al, 1984). Ces composéspourraient soit avoir une activitéauxinique, soit présenter une activitédifférente ainsi qu’un niveau de spécificitéplus élévé que celui de l’auxine.
En accord avec l’implication de CgAUX1dans le mécanisme d’infection, l’utilisationde l’inhibiteur spécifique du transportd’influx d’auxine 1-NOA entraine unretard dans l’apparition des prénodules etdes nodules. Ce retard dans la nodulationest diminué par l’ajout de l’auxinelipophile NAA ce qui montre qu’il s’agitd’un effet spécifique sur l’influx d’auxine.Cependant, l’utilisation du 1-NOAprésente certaines limites : la pénétrationdu 1-NOA dans les tissus internes de laracine de C. glauca est probablementassez limitée. De plus, le 1-NOA est uninhibiteur compétitif, il y a donc de forteschances que sa concentration ne soit passuffisamment élevée pour inhiber letransport d’influx d’auxine dans les tissusles plus internes de la racine. De la mêmemanière, la forte production d’auxine parFrankia dans un site confiné, comme lazone de courbure du poil racinaire,pourrait limiter l’effet du 1-NOA.Autrement dit, son action ne permetprobablement que de réduire l’efficacitédu processus d’infection. Cela peut suffireà expliquer que le traitement avec du 1-NOA ne permet pas d’empêcher l’infectionmais entraine juste un retard de la mise enplace de la symbiose. La technique d’ARNinterférent a récemment été développéechez C. glauca dans notre laboratoire.L’extinction spécifique de CgAUX1permettra de préciser dans quelle mesurele transport d’influx d’auxine est impliquédans le mécanisme d’infection par Frankiaau cours de la symbiose actinorhizienne.
2) CgAUX1 pourrait créer un puitsd’auxine au site d’infectionL’infection par la bactérie Frankia est
associée à une sur-expression dans lesystème vasculaire en regard du sited’infection (Figure 35). La production d’untransporteur d’influx d’auxine peutpermettre de modifier les flux d’auxine ausite d’infection. Chez Arabidopsis,l’expression d’AtAUX1 permet d’initier letransport post-phloèmien de l’auxine versl’apex de la racine (Swarup et al, 2001). Defaçon similaire, la sur-expression deCgAUX1 dans le tissu vasculaire pourraitpermettre de créer un puits d’auxine etd’augmenter localement sa concentration.Une augmentation de la concentration enauxine est d’ailleurs observée au sited’infection chez les Légumineuses(Pacios-Bras et al, 2003). Cependant, cetteaccumulation est liée à une inhibition dutransport d’efflux (Mathesius et al, 1998).Aucune information sur le transportd’efflux d’auxine au cours de la symbioseactinorhizienne n’est disponible. Desétudes par hybridation in situ surl’ensemble des gènes LAX de M. truncatulaont montré que le transport d’influxd’auxine n’est pas augmenté dans lestissus vasculaires au site d’infection (deBilly et al, 2001). Néanmoins, l’utilisationd’une sonde unique pour détecter lesARNm de cinq gènes par hybridation insitu ne permet peut-être pas d’obtenir unniveau de détection suffisamment élevé.Quoi qu’il en soit, l’augmentation de laconcentration en auxine pourrait êtreresponsable des divisions cellulaires dansle cortex pour former le prénodule et dansle péricycle pour former le nodule chez C.glauca. En effet, l’expression de CgAUX1est clairement visible dans ces deux tissus.
3) CgAUX1 n’est pas impliqué dansl’organisation du primordiumnodulaireLe nodule actinorhizien est considéré
comme une racine latérale modifiée(Pawlowski et Bisseling, 1996). Le profild’expression de CgLAX3 très semblableentre ces deux organes semble confirmerce postulat (Figures 34 et 36). Néanmoins,CgAUX1 est exprimé dans le primordium

41
racinaire mais pas dans le primordiumnodulaire (Figure 35). Ces résultatsindiquent que ces deux organes ont desdivergences dans leurs programmesdéveloppementaux. Ceci est en accordavec d’autres observations qui montrentque l’expression hétérologue depromoteurs comme le 35S ou AtUBQ1 ontdes profils d’expression différents dans leprimordium de racine et de nodule chezles Casuarinacées (Obertello et al, 2005).
Plusieurs hypothèses peuvent expliquerla différence de profil d’expression deCgAUX1. Soit le primordium nodulaire nenécessite pas de transporteur d’influxd’auxine pour son organisation, soit unautre gène que CgAUX1 remplit cettefonction. Le transport d’auxine dans leprimordium de racine latérale est associé àla création d’un gradient d’auxine dont lemaximum se trouve à l’apex (Benková etal, 2003). Néanmoins, ce sontessentiellement les transporteurs d’effluxqui assument ce rôle. L’absence detransporteur d’influx dans le primordiumpourrait donc ne pas perturber les fluxd’auxine nécessaires à son organisation.D’ailleurs, le mutant aux1 d’A. thaliana estperturbé dans l’initiation de la racinelatérale mais pas dans l’organisation du
primordium (Marchant et al, 2002).Malheureusement, l’analyse du profild’expression de CgLAX3 ne nous a paspermis de déterminer s’il est exprimé ounon dans le primordium nodulaire. Il estdonc possible qu’un autre gène AUX-LAXque CgAUX1 soit exprimé dans leprimordium de nodule.
L’utilisation d’une technique detransformation génétique plus rapidebasée sur la production de chevelusracinaires (hairy root) par Agrobacteriumrhizogenes permettra dans un avenirproche de générer des racines de C.glauca contenant la constructionProCgLAX3:GUS. Cette technique estdésormais maitrisée dans notrelaboratoire. L’analyse de l’expression deCgLAX3 révelera si ce gène est exprimédans le primordium de nodule. Afin depréciser le rôle de ce gène, l’extinction deson expression par ARN interférent pourraêtre envisagée. De plus, le séquençage encours d’un grand nombre d’EST par notreéquipe dans le cadre d’un projetGénoscope permettra éventuellementd’identifier un nouveau gène de la familleAUX-LAX qui aurait pu échapper à nosdifférentes approches pour identifier tousles gènes de la famille. Le cas échéant, le
clonage et la caractérisation de ce gèneseraient initiés afin d’obtenir des
informations similaires à celles qui ont étéobtenues pour CgAUX1 et CgLAX3.

42
PARTIE III - CIBLES DE LA SIGNALISATION AUXINIQUE DEPENDANTEDU TRANSPORT D’INFLUX
I. IntroductionLes résultats que nous avons obtenus
suggèrent que le transport d’influxd’auxine est impliqué dans le mécanismed’infection par Frankia au cours de lasymbiose actinorhizienne. Néanmoins, iln’existe aucune information sur les ciblesde la signalisation auxinique au cours del’infection des plantes actinorhiziennes.Chez Arabidopsis thaliana, aucune cible dela signalisation auxinique dépendante dugène AtAUX1 n’est connue. Afind’identifier ces cibles au cours de lasymbiose actinorhizienne, nous avonsdécidé de collaborer à l’identification deces mécanismes chez la plante modèle.L’identification récente du mutant lax3 parle laboratoire du professeur MalcolmBennett (Université de Nottingham,Royaume-Uni) a permis de mettre enévidence de nouveaux mécanismesimpliqués dans l’émergence de la racinelatérale. Nous nous sommes impliquésdans l’identification des mécanismessitués en aval de l’action du gène AtLAX3au cours de ce processus. Dans cetteintroduction, nous présenterons donc lesrésultats obtenus par le laboratoire de M.Bennett sur la caractérisation du mutantlax3 puis dans une seconde partie lestravaux réalisés dans le cadre de cettethèse sur l’identification des cibles de lasignalisation auxinique dépendanted’AtLAX3.
1) AtLAX3 est impliqué dansl’émergence de la racine latéraleTrois homologues du gène AtAUX1 ont
été identifiés chez A. thaliana : AtLAX1,AtLAX2 et AtLAX3. L’identification delignées d’insertion pour chacun de cesgènes a permis de mettre en évidence unphénotype de réduction du nombre de
racines latérales pour le mutant lax3 alorsque les mutants lax1 et lax2 n’ont pas dephénotype racinaire. Le mutant lax3présente une réduction d’environ 40% dunombre de racines latérales émergées parcomparaison avec le type sauvage (Figure40a-e). Ce phénotype est comparable àcelui du mutant aux1 (Marchant et al,2002). Contrairement au mutant aux1 quiprésente environ moitié moins deprimordia que le sauvage, le mutant lax3possède trois fois plus de primordia que lesauvage. Le phénotype du mutant lax3 nepeut donc pas s’expliquer par un défautd’initiation comme c’est le cas pour aux1.À l’inverse, la mutation lax3 arrête laprogression du primordium car la majoritédes primordia sont bloqués au stade I(Figure 40f).
2) AtLAX3 est exprimé dans lescellules en face du primordiumLa protéine LAX3 est présente dans la
stèle et les quelques cellules du cortex etde l’épiderme situées en face duprimordium de racine latérale (Figure 41).Ce profil d’expression est distinct de celuid’AtAUX1 (Marchant et al, 2002) et
Figure 40. Phénotype du mutant lax3Nombre de racines latérales chez le mutant lax3, aux1-21,le double mutant lax3aux1-21 comparé au sauvage Col012 jours après germination (a). Jeunes plants sauvagesCol0 (b), mutants lax3 (c), mutants aux1-21 (d) et doublemutants lax3aux1-21 (e) 12 jours après germination.Nombre de primordia pour chaque stade dedéveloppement 8 jours après germination (f). Les valeursstatistiquement différentes du témoin (test de StudentP<0,05) sont indiquées par un astérisque.

43
explique l’additivité des phénotypes aux1et lax3. En effet le double mutantaux1lax3 ne produit pas de racineslatérales pendant les 14 premiers joursaprès germination (Figure 40e). De plus,ce profil d’expression indique que LAX3permet le développement du primordiumde manière indirecte à partir de la stèle etdes cellules corticales et épidermiques.
La production de la protéine LAX3uniquement dans la stèle en utilisant lepromoteur du gène SHORTROOT(Helariutta et al, 2000) ne permet pas derestaurer le phénotype sauvage desmutants lax3 (donnée non montrée – voirAnnexes). Par contre, l’expression de LAX3sous contrôle de son propre promoteurrestaure le phénotype sauvage. De plus,l’utilisation d’un fusion fonctionnelleLAX3:YFP montre que la protéine LAX3n’est pas localisée à la surface des cellulesde la stèle alors que la localisation de LAX3est bien membranaire dans les cellulescorticales et épidermiques.
Cela montre que l’expression de LAX3dans les cellules du cortex et del’épiderme est nécessaire pour permettrede restaurer un phénotype racinairesauvage.
3) AtLAX3 est inductible par l’auxineDes analyses par RT-PCR quantitative
ont montré que LAX3 est inductible parl’auxine (Figure 42E). Ces données ont étéprécisées par l’utilisation d’une fusionLAX3:YFP sous dépendance du promoteurde LAX3 (Figure 42A et B). En effet,
l’induction par l’auxine est spécifique ducortex et, dans une moindre mesure, del’épiderme. Cette inductibilité par l’auxineest dépendante du gène SOLITARY ROOT(SLR1/IAA14 ; Figure 42C à E) qui estégalement exprimé dans le cortex etl’épiderme de manière assez faible (Fukakiet al, 2002). En effet, la mutation de gainde fonction slr1 bloque l’inductibilité deLAX3 par l’auxine.
L’auxine s’accummule dans l’apex duprimordium de racine latérale au cours deson développement (Benková et al, 2003).L’utilisation du gène rapporteur DR5 apermis de mettre en évidence une réponse
iFigure 41. Profil d’expressiond’AtLAX3Expression d’AtLAX3déterminée par l’utilisation de laconstruction ProAtLAX3:GUS.AtLAX3 est exprimé dans lesystème vasculaire (a) jusqu’à lazone d’élongation (b).Expression d’AtLAX3 au niveaud’une coupe transversale deracine (c) et d’une jeune racinelatérale (d). AtLAX3 n’est pasexprimé dans le primordium deracine latérale mais dans lestissus adjacents : stade I (e),stade II (f), stade V (g), stade VII(h) et après émergence (fusionLAX3:YFP – i). Barres : 25µm.
Figure 42. AtLAX3 est induit par l’auxine de façondépendante de SLR1/IAA14Expression d’AtLAX3 déterminée par fusion LAX3:YFPaprès traitement avec 10–8M d’IAA (A) ou 10-7M d’IAA (B).Expression du marqueur DR5:GUS dans le cortex etl’épiderme après traitement avec 10-5M de NAA chez lesauvage Col0 (C) et chez le mutant slr1 (D). Inductiond’AtLAX3 par l’auxine (10-5M de NAA) déterminé par RT-PCR quantitative chez Col0 ou chez le mutant slr1 (E).

44
dans les tissus situés en face duprimordium, suggérant un passage del’auxine contenue dans le primordium versles tissus adjacents. Ce signal estexclusivement observé dans les cellules del’endoderme, du cortex et de l’épidermesituées en regard du primordium.
L’émergence des primordia de racinelatérale est augmentée chez le mutantpin2. La mutation du gène PIN2 perturbele transport basipète d’auxine dans laracine primaire (Rashotte et al, 2000).L’analyse du profil d’expression de PIN2suggère que PIN2 permet le transportbasipète dans le méristème apical et deracine latéral (Figure 43a). En accord avecces observations, le marqueur DR5 montreune plus forte accummulation d’auxinedans l’apex des primordia de racineslatérales chez le mutant pin2 (Figure 43b àe). Cette plus forte accumulation d’auxineest associée à une augmentation del’émergence chez le mutant pin2.
4) ConclusionLe profil d’expression de LAX3 semble
donc résulter d’une induction par l’auxineen provenance des primordia, ce quiexplique la spécificité d’expression deLAX3 dans les quelques cellules en regarddu primordium. Nous avons cherché àidentifier les gènes cibles de lasignalisation auxinique dans ces cellulesdu cortex et de l’épiderme impliquéesdans l’émergence de la racine latérale. Lesgènes impliqués dans le remodelage de laparoi sont des candidats potentiels carcertains d’entre eux sont exprimés lors del’émergence. Par la suite, des homologuesde ces gènes seront identifiés chez C.
glauca. L’objectif est de découvrir lesgènes induits en réponse à un signalauxinique sous dépendance d’untransporteur d’influx dans le cas del’infection par Frankia au cours de lasymbiose actinorhizienne.
II. AtLAX3 permet leremodelage de la paroidans les cellules del’épiderme et du cortexau cours de l’émergence
Pour identifier les cibles de lasignalisation auxinique dans les cellulesqui expriment LAX3, nous avons utilisédeux approches. Tout d’abord uneapproche gène candidat a été menée enidentifiant un gène ayant le même profild’expression que LAX3. Ensuite, desanalyses transcriptomiques nous ontfourni une liste de gènes induits parl’auxine et exprimés dans le cortex etl’épiderme.
1) L’induction d’AIR3 par l’auxinedépend de LAX3L’émergence de la racine latérale
implique la séparation des cellules situéesen face du primordium au cours de saprogression à travers ces tissus. Cescellules restent intactes et se séparententre elles au niveau de leur lamellemoyenne (Laskowski et al, 2006). Le gèneAIR3 code une protéase de type subtilisineet est exprimé dans les cellules du cortexet de l’endoderme situées en face duprimordium de racine latérale (Neuteboomet al, 1999b). De plus, l’expression d’AIR3est induite par l’auxine (Neuteboom et al,1999b). Ce gène présente donc le mêmeprofil d’expression qu’AtLAX3 et constitueun candidat idéal car il pourrait êtreimpliqué dans le remodelage de la paroiau cours du processus d’émergence. Nousavons donc testé si cette induction parl’auxine est dépendante de LAX3. Pourcela, la construction ProAIR3 :GUS a étéintroduite chez le mutant lax3 et chez lesauvage (Col0). Les analyses ont étéréalisées sur 3 lignées T2 pour lax3 et 5lignées T2 pour Col0. De manièreintéressante, la mutation lax3 entraine une
Figure 43. PIN2 permet le transport basipète del’auxine dans le primordiumExpression de PIN2 dans le primordium de racine latéraldéterminée par fusion PIN2:GFP (a). Maximum d’auxine àl’apex du primordium démontré par l’utilisation dumarqueur DR5:GUS (b) ou DR5:GFP (d). Ce maximum estplus élevé chez le mutant pin2 (c et e).
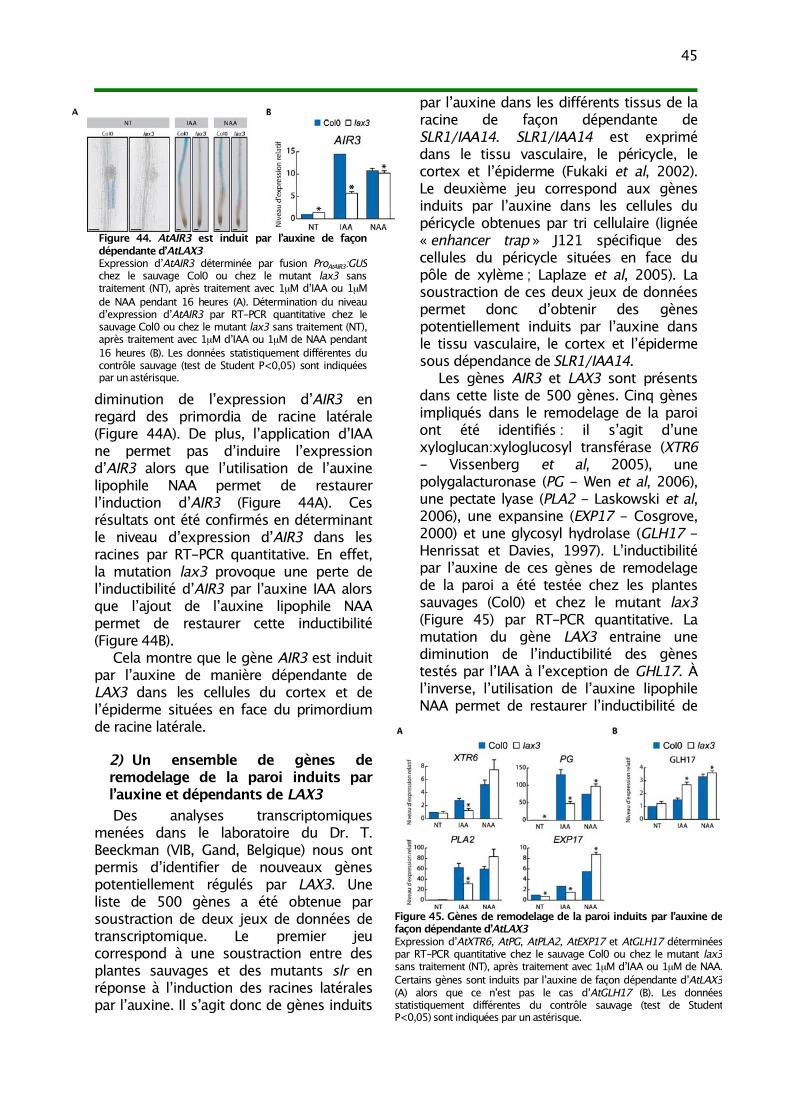
45
diminution de l’expression d’AIR3 enregard des primordia de racine latérale(Figure 44A). De plus, l’application d’IAAne permet pas d’induire l’expressiond’AIR3 alors que l’utilisation de l’auxinelipophile NAA permet de restaurerl’induction d’AIR3 (Figure 44A). Cesrésultats ont été confirmés en déterminantle niveau d’expression d’AIR3 dans lesracines par RT-PCR quantitative. En effet,la mutation lax3 provoque une perte del’inductibilité d’AIR3 par l’auxine IAA alorsque l’ajout de l’auxine lipophile NAApermet de restaurer cette inductibilité(Figure 44B).
Cela montre que le gène AIR3 est induitpar l’auxine de manière dépendante deLAX3 dans les cellules du cortex et del’épiderme situées en face du primordiumde racine latérale.
2) Un ensemble de gènes deremodelage de la paroi induits parl’auxine et dépendants de LAX3Des analyses transcriptomiques
menées dans le laboratoire du Dr. T.Beeckman (VIB, Gand, Belgique) nous ontpermis d’identifier de nouveaux gènespotentiellement régulés par LAX3. Uneliste de 500 gènes a été obtenue parsoustraction de deux jeux de données detranscriptomique. Le premier jeucorrespond à une soustraction entre desplantes sauvages et des mutants slr enréponse à l’induction des racines latéralespar l’auxine. Il s’agit donc de gènes induits
par l’auxine dans les différents tissus de laracine de façon dépendante deSLR1/IAA14. SLR1/IAA14 est exprimédans le tissu vasculaire, le péricycle, lecortex et l’épiderme (Fukaki et al, 2002).Le deuxième jeu correspond aux gènesinduits par l’auxine dans les cellules dupéricycle obtenues par tri cellulaire (lignée« enhancer trap » J121 spécifique descellules du péricycle situées en face dupôle de xylème ; Laplaze et al, 2005). Lasoustraction de ces deux jeux de donnéespermet donc d’obtenir des gènespotentiellement induits par l’auxine dansle tissu vasculaire, le cortex et l’épidermesous dépendance de SLR1/IAA14.
Les gènes AIR3 et LAX3 sont présentsdans cette liste de 500 gènes. Cinq gènesimpliqués dans le remodelage de la paroiont été identifiés : il s’agit d’unexyloglucan:xyloglucosyl transférase (XTR6- Vissenberg et al, 2005), unepolygalacturonase (PG - Wen et al, 2006),une pectate lyase (PLA2 - Laskowski et al,2006), une expansine (EXP17 - Cosgrove,2000) et une glycosyl hydrolase (GLH17 -Henrissat et Davies, 1997). L’inductibilitépar l’auxine de ces gènes de remodelagede la paroi a été testée chez les plantessauvages (Col0) et chez le mutant lax3(Figure 45) par RT-PCR quantitative. Lamutation du gène LAX3 entraine unediminution de l’inductibilité des gènestestés par l’IAA à l’exception de GHL17. Àl’inverse, l’utilisation de l’auxine lipophileNAA permet de restaurer l’inductibilité de
Figure 44. AtAIR3 est induit par l’auxine de façondépendante d’AtLAX3Expression d’AtAIR3 déterminée par fusion ProAtAIR3:GUSchez le sauvage Col0 ou chez le mutant lax3 sanstraitement (NT), après traitement avec 1µM d’IAA ou 1µMde NAA pendant 16 heures (A). Détermination du niveaud’expression d’AtAIR3 par RT-PCR quantitative chez lesauvage Col0 ou chez le mutant lax3 sans traitement (NT),après traitement avec 1µM d’IAA ou 1µM de NAA pendant16 heures (B). Les données statistiquement différentes ducontrôle sauvage (test de Student P<0,05) sont indiquéespar un astérisque.
Figure 45. Gènes de remodelage de la paroi induits par l’auxine defaçon dépendante d’AtLAX3Expression d’AtXTR6, AtPG, AtPLA2, AtEXP17 et AtGLH17 déterminéespar RT-PCR quantitative chez le sauvage Col0 ou chez le mutant lax3sans traitement (NT), après traitement avec 1µM d’IAA ou 1µM de NAA.Certains gènes sont induits par l’auxine de façon dépendante d’AtLAX3(A) alors que ce n’est pas le cas d’AtGLH17 (B). Les donnéesstatistiquement différentes du contrôle sauvage (test de StudentP<0,05) sont indiquées par un astérisque.

46
ces gènes de remodelage de la paroi. Afinde confirmer ces résultats, desconstructions promoteur:GUS ont étéréalisées pour ces gènes (à l’exception deGLH17). Pour les quatres gènes (XTR6, PG,PLA2 et EXP17), une séquence de 1000pdb située en amont de l’ATG a étéamplifiée par PCR et clonée dans le vecteurpBI101.3 en amont du gène rapporteurGUS. La transformation génétique deplantes sauvages (Col0) et mutantes (lax3)d’A. thaliana est en cours. L’objectif est devérifier que ces gènes sont bien exprimésdans les mêmes tissus que LAX3 au coursde l’émergence de la racine latérale et deconfirmer que ce profil d’expression estdépendant de LAX3.
3) Modèle d’action d’AtLAX3 dansl’émergence de la racine latéraleLa libération d’auxine par le
primordium racinaire permet l’inductiond’AtLAX3 dans le cortex juste en face duprimordium (Figure 46). L’expression deLAX3 dans ce tissu permet la productiond’un certain nombre de gènes deremodelage de la paroi. La simplediffusion de l’auxine à partir duprimordium entrainerait un gradientd’expression des gènes de remodelage dela paroi à partir de la zone de productiond’auxine (apex du primordium). Or cen’est pas ce que l’on observe car seules lescellules directement en contact avec leprimordium expriment ces gènes. Aucontraire, l’induction de LAX3 par l’auxine
provoque la mise en place d’une boucle derétrocontrôle positif car la production deplus de transporteurs en réponse àl’auxine augmente l’influx d’auxine dansla cellule. Cela explique la réponse de type« tout ou rien » observée dans les cellulescorticales et épidermiques adjacentes duprimordium qui expriment AtLAX3 et lesgènes de remodelage de la paroi (Figure41i et 44A). Cette boucle d’amplificationpermet de créer localement un puitsd’auxine dans les quelques cellules qui sesituent à proximité immédiate duprimordium et de ce fait limite la diffusionde l’auxine dans les cellules adjacentes.Un certain nombre de gènes facilitant leremodelage de la paroi sont induits etpermettent le passage du primordium àtravers les cellules du cortex puis del’épiderme (Figure 46). La séparation descellules est facilitée par la productiond’enzymes telles que les pectate lyases(Marín-Rodríguez et al, 2002) et lespolygalacturonases (Wen et al, 2006). Cesenzymes sont paticulièrement importantesau cours de l’émergence car les cellulesqui entourent le primordium sont richesen pectines non-méthylées (Laskowski etal, 2006) qui sont le substrat des pectatelyases et des polygalacturonases.
Le mécanisme présenté ici permet doncà la fois d’amplifier le faible signalauxinique provenant du primordium et dele restreindre aux cellules qui sontdirectement en contact avec celui-ci. Eneffet, une large diffusion de l’auxine
Figure 46. Modèle du rôle d’AtLAX3 au cours de l’émergenceL’auxine qui proviendrait des cellules du primordium induit l’expression d’un jeu de gènes de remodelage de la paroi (CWR)dans l’endoderme en levant l’inhibition exercée par SHY2/IAA3 (A). Une faible quantité d’auxine parvient jusqu’aux cellules ducortex par diffusion et induit l’expression d’AtLAX3 en levant l’inhibition exercée par SLR/IAA14 (B). AtLAX3 permet l’entrée deplus d’auxine dans la cellule ce qui initie une boucle de rétrocontrôle positif et permet l’induction d’un jeu de gènes deremodelage de la paroi (C). La progression du primordium est facilitée par le remodelage de la paroi et la production d’enzymesde remodelage de la paroi dans l’épiderme est permise par une nouvelle amplification du signal auxinique (D). Couleur bleue :niveau d’expression du marqueur DR5:GUS, P : péricycle, End : endoderme, C : cortex, Epi : épiderme.

47
provoquerait le remodelage de la paroidans une zone plus large autour duprimordium et pourrait par exemplefaciliter l’infection par des bactériespathogènes. Cela expliquerait égalementpourquoi l’expression de LAX3 dansl’épiderme est moins facilement induitepar l’auxine que dans le cortex.
III. Influx d’auxine etremodelage de la paroiau cours de l’infectionpar Frankia
Les résultats obtenus sur le rôled’AtLAX3 chez Arabidopsis suggèrent queCgAUX1 pourrait être impliqué dansl’amplification du signal symbiotiqueactinorhizien et la restriction de ce signaldans les cellules infectées. De plus, cetteamplification pourrait permettrel’expression de gènes de remodelage de laparoi. En effet, le processus d’infection parFrankia au cours de la symbioseactinorhizienne est associé au remodelagede la paroi pour la création du cordond’infection (Berg, 1999a ; Berry et al, 1986)et des ponts cytoplasmiques (Berg,1999b). De la même manière, leremodelage de la paroi est impliqué dansla mise en place de la symbioseRhizobium-Légumineuses (Brewin, 2004).La production des facteurs Nod par labactérie provoque la courbure du poilracinaire qui nécessite une modification dela croissance de la paroi et le cordond’infection est une structure tubulaireformée de matériel pariétal qui pénètredans la cellule (Brewin, 2004). Néanmoins,les mécanismes moléculaires associés àces modifications sont inconnus chez lesplantes actinorhiziennes. Chez lesLégumineuses, de nombreuses nodulinesprécoces sont des glycoprotéines riches enhydroxyproline (HRGP) dont le rôle dans lamodification de la paroi est suspecté(Cassab, 1998). Mais dans ce cas, letransport d’influx d’auxine ne semble pasêtre impliqué dans l’expression de cesgènes car le transport d’influx d’auxinen’est pas associé avec l’infection parRhizobium (de Billy et al, 2001).
Nous avons donc cherché à identifierdes cibles potentielles de la signalisationauxinique au cours de l’infection de lacellule végétale par Frankia par homologieavec les gènes cibles de la signalisationdépendante d’AtLAX3. Un gène nomméCg12 et codant une protéase de typesubtilisine appartenant à la même famillequ’AtAIR3 est exprimé de manièrespécifique dans les cellules infectées parFrankia au cours de la symbioseactinorhizienne chez C. glauca(Svistoonoff et al, 2003 ; Svistoonoff,2003). Selon notre modèle, il serait doncpossible que Cg12 soit induit par l’auxinequi entre dans la cellule grâce autransporteur CgAUX1. Afin d’élargir cetteanalyse, nous avons également identifiéd’autres gènes de remodelage de la paroidans nos banques EST et nous avonsdéterminé par RT-PCR quantitative i) s’ilsétaient induits par l’auxine et ii) s’ilsétaient associés au processus d’infection.
1) Régulation de Cg12 par un signalauxiniqueNous avons déterminé si l’auxine induit
l’expression du gène Cg12 en utilisant laRT-PCR quantitative. Aucune induction dugène n’a pu être observée après untraitement de 24 heures avec 1µM ou10µM d’IAA, NAA ou PAA (données nonmontrées). Ces résultats confirment desétudes précédentes de l’expressionconférée par la construction ProCg12 :GUSchez C. glauca (Svistoonoff, 2003).L’auxine seule n’est donc pas responsabledu profil d’expression de Cg12 au coursde l’infection.
Afin de déterminer si une diminution del’entrée d’auxine dans la cellule a un effetsur l’expression de Cg12, des plants de C.glauca ont été inoculés par la bactérieFrankia en présence de 50 µM del’inhibiteur de transport d’influx 1-NOA.Nous avons ensuite déterminé le niveaud’expression de Cg12 dans les nodulesmatures âgés de 24 jours chez les plantestraitées avec du 1-NOA par comparaisonaux nodules du même âge chez desplantes non traitées. Les résultatsmontrent que l’expression de Cg12 estlargement diminuée par le traitement au
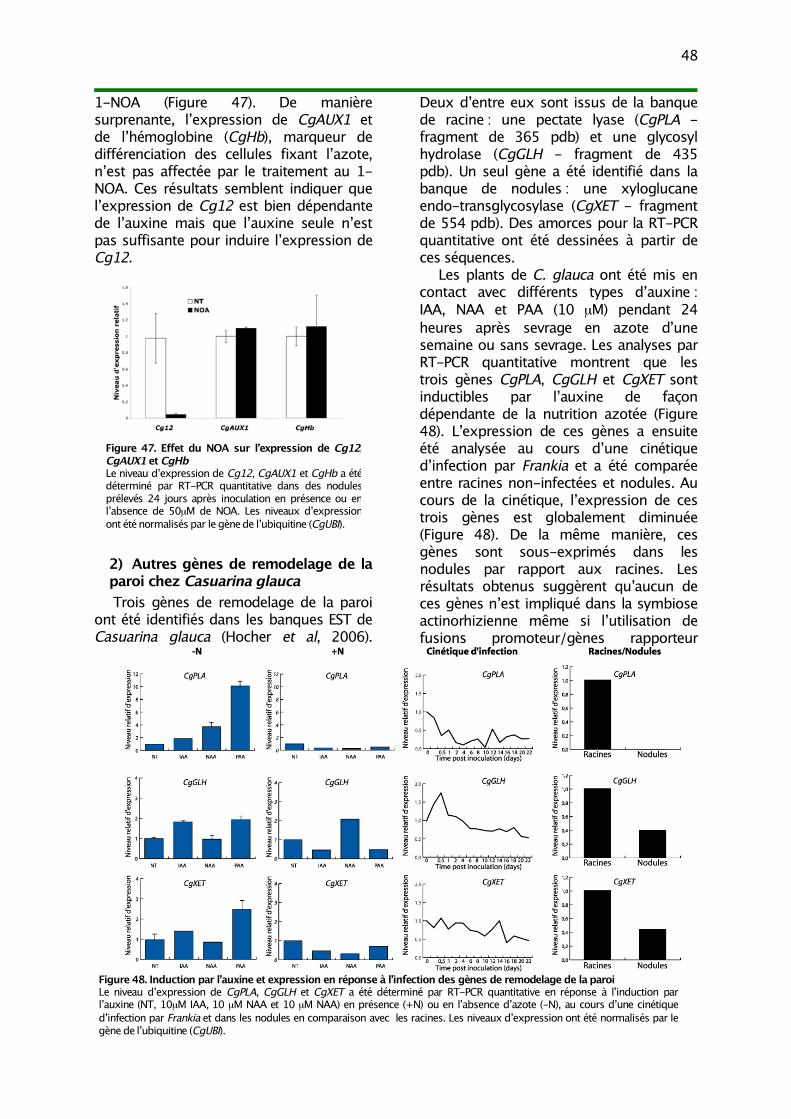
48
1-NOA (Figure 47). De manièresurprenante, l’expression de CgAUX1 etde l’hémoglobine (CgHb), marqueur dedifférenciation des cellules fixant l’azote,n’est pas affectée par le traitement au 1-NOA. Ces résultats semblent indiquer quel’expression de Cg12 est bien dépendantede l’auxine mais que l’auxine seule n’estpas suffisante pour induire l’expression deCg12.
2) Autres gènes de remodelage de laparoi chez Casuarina glaucaTrois gènes de remodelage de la paroi
ont été identifiés dans les banques EST deCasuarina glauca (Hocher et al, 2006).
Deux d’entre eux sont issus de la banquede racine : une pectate lyase (CgPLA -fragment de 365 pdb) et une glycosylhydrolase (CgGLH - fragment de 435pdb). Un seul gène a été identifié dans labanque de nodules : une xyloglucaneendo-transglycosylase (CgXET - fragmentde 554 pdb). Des amorces pour la RT-PCRquantitative ont été dessinées à partir deces séquences.
Les plants de C. glauca ont été mis encontact avec différents types d’auxine :IAA, NAA et PAA (10 µM) pendant 24heures après sevrage en azote d’unesemaine ou sans sevrage. Les analyses parRT-PCR quantitative montrent que lestrois gènes CgPLA, CgGLH et CgXET sontinductibles par l’auxine de façondépendante de la nutrition azotée (Figure48). L’expression de ces gènes a ensuiteété analysée au cours d’une cinétiqued’infection par Frankia et a été comparéeentre racines non-infectées et nodules. Aucours de la cinétique, l’expression de cestrois gènes est globalement diminuée(Figure 48). De la même manière, cesgènes sont sous-exprimés dans lesnodules par rapport aux racines. Lesrésultats obtenus suggèrent qu’aucun deces gènes n’est impliqué dans la symbioseactinorhizienne même si l’utilisation defusions promoteur/gènes rapporteur
Figure 47. Effet du NOA sur l’expression de Cg12,CgAUX1 et CgHbLe niveau d’expression de Cg12, CgAUX1 et CgHb a étédéterminé par RT-PCR quantitative dans des nodulesprélevés 24 jours après inoculation en présence ou enl’absence de 50µM de NOA. Les niveaux d’expressionont été normalisés par le gène de l’ubiquitine (CgUBI).
Figure 48. Induction par l’auxine et expression en réponse à l’infection des gènes de remodelage de la paroiLe niveau d’expression de CgPLA, CgGLH et CgXET a été déterminé par RT-PCR quantitative en réponse à l’induction parl’auxine (NT, 10µM IAA, 10 µM NAA et 10 µM NAA) en présence (+N) ou en l’absence d’azote (-N), au cours d’une cinétiqued’infection par Frankia et dans les nodules en comparaison avec les racines. Les niveaux d’expression ont été normalisés par legène de l’ubiquitine (CgUBI).

49
permettrait de confirmer cette approche.Seuls trois gènes impliqués dans le
remodelage de la paroi ont été identifiésdans les banques EST. À l’heure actuelle,ces banques contiennent environ 1 000séquences de gènes exprimés dans lesnodules et autant dans les racines, ainsiqu’environ 400 correspondant à unebanque soustractive entre des ADNc issusde racines non infectées et des racinessept jours après infection. Le séquençagede 10 000 EST supplémentaires exprimésdans les nodules et autant dans les racinesainsi que 2 000 EST de banquessoustractives (correspondant à deux etquatre jours après infection) devraitpermettre d’identifier de nombreuxnouveaux candidats.
IV. DiscussionL’analyse du rôle d’AtLAX3 au cours de
l’émergence a permis d’identifier descibles de la signalisation auxinique liée àl’influx d’auxine au cours de l’émergence.L’infection de la cellule végétale parFrankia entraine un remodelage de la paroinécessaire pour la mise en place ducordon d’infection (Berg, 1999a ; Berry etal, 1986) et des ponts cytoplasmiques(Berg, 1999b). Or nos résultats montrentque l’influx d’auxine est étroitement lié àl’infection. Sur la base du modèle proposépour l’émergence de la racine latérale,nous avons donc émis l’hypothèse quel’influx d’auxine, probablement d’originebactérienne, permet l’expression de gènesde remodelage de la paroi au cours del’infection par un mécanismed’amplification du signal symbiotique.
1) Amplification du signal auxiniquepar l’influx d’auxineSur la base des résultats obtenus, nous
proposons un modèle de l’action dutransport d’influx d’auxine au cours del’infection par Frankia (Figure 49). Il estcouramment admis qu’un signalsymbiotique de nature inconnue estproduit par Frankia (Pawlowski etSirrenberg, 2003). Ce signal permetd’initier les premières étapes de l’infectionet probablement d’induire l’expression deCgAUX1. Le transporteur CgAUX1, en
favorisant l’influx d’auxine bactérienne,améliore le processus d’infection et meten place une boucle de rétrocontrôlepositif. Ce mécanisme permettrait ausignal auxinique d’atteindre un niveausuffisant pour induire les gènes cibles,dont le gène Cg12 impliqué dans leremodelage de la paroi. Par homologieavec l’émergence de la racine latérale, ilest probable qu’une amplification dusignal symbiotique dans la cellule infectéepermet d’obtenir une réponse de type« tout ou rien » uniquement dans cescellules. En effet, dans un tel système, leniveau d’auxine suffisant pour entrainerune réponse n’est atteint que dans lescellules qui expriment le transporteurd’influx CgAUX1.
Si notre modèle est correct,l’expression des gènes de remodelage dela paroi nécessite à la fois un signalsymbiotique et un apport d’auxine au-delà d’un certain seuil (Figure 49). En effet,même si l’auxine seule ne peut induirel’expression de Cg12, l’inhibition dutransport d’influx d’auxine par le 1-NOA apour conséquence de réduire son niveaud’expression d’un facteur 20 environ sansaffecter l’expression de CgAUX1. Celamontre que l’induction de Cg12 au coursde l’infection est bien, du moins en partie,dépendante de l’auxine. L’amplification dusignal spécifique par l’entrée d’auxinedans la cellule correspond donc à uneaugmentation de l’expression de Cg12
Figure 49. Modèle du rôle de CgAUX1 au cours del’infection par FrankiaLa perception d’un facteur de nodulation par Frankiapermet d’initier l’infection et d’induire le gène CgAUX1. Laproduction du transporteur d’influx d’auxine permetl’entrée d’auxine (d’origine bactérienne ?) qui amplifie lesignal symbiotique et permet d’augmenter l’expression degènes de remodelage de la paroi comme Cg12.

50
d’un facteur 20. La valeur réelle pourraits’avérer encore plus élevée que cettevaleur théorique car l’utilisation de 1-NOAne permet probablement pas d’inhibertotalement le transport d’influx. Latechnique d’ARN interférent permettrait dedéterminer la diminution d’expression deCg12 en l’absence totale de transportd’influx et donc de quantifierl’amplification liée à l’entrée d’auxine.
Par ailleurs, l’identification d’autresgènes de remodelage de la paroiprésentant un profil d’expressionidentique à celui de Cg12 s’avèreindispensable pour confirmer cesobservations. Le séquençage en cours denombreuses EST issues de nodules etracines (deux jours et quatre jours après
inoculation) devrait permettred’identifier de nouveaux candidats. Parmiceux-ci, les gènes qui ne sont pas induitspar l’auxine seule mais sont exprimés enréponse à l’infection et dont l’expressionest réduite en réponse au traitement par le1-NOA pourront être identifiés par RT-PCR. Le clonage des séquencespromotrices de ces gènes et l’analyse deleur profil d’expression par fusionpromoteur:GUS permettra de déterminer sices gènes sont bien liés à l’infection de lacellule végétale par Frankia. L’insertion deces constructions dans les plantes oùl’expression de CgAUX1 a été éteinte parARN interférent permettra de déterminer sice profil d’expression est lié à l’action deCgAUX1. Sur la base des résultats obtenuschez A. thaliana, il est probable que leremodelage de la paroi au cours del’infection implique également plusieursgènes.
2) Nature du signal symbiotiqueLe mécanisme d’amplification serait
d’autant plus efficace si le signalsymbiotique était un composé indoliquecar il jouerait alors à la fois le rôled’amplificateur et de signal spécifique. Eneffet, il n’est pas improbable que CgAUX1puisse prendre en charge des composésayant une structure proche de l’auxineIAA. Dans ce cas, la présence de motifs surun noyau indolique pourrait conférer à cesmolécules une capacité à se lier avec unrécepteur spécifique. Bien que purementspéculatif, ce modèle est toutefoisenvisageable car la nature chimique dessignaux symbiotiques actinorhiziens et
des facteurs Nod est différente (Cérémonieet al, 1999). Les tentatives d’induction desgènes Cg12 et CgAUX1 avec dusurnageant de Frankia n’ont donné aucunrésultat. Néanmoins, toutes cesexpériences ont été réalisées avec dusurnageant de bactéries qui n’avaient pasété mises en contact avec la plante hôte. Ila été montré que le facteur dedéformation des poils absorbants del’aulne est produit avec ou sans inductionde la bactérie par des exsudats racinaires(van Ghelue et al, 1997). Néanmoins, il estprobable que le facteur de déformationdes poils racinaires ne soit pas le signalsymbiotique car la déformation du poil estun processus moins spécifique chez lesplantes actinorhiziennes que chez lesLégumineuses. Les facteurs Nod deRhizobium ne sont pas produits demanière constitutive mais en réponse auxflavonoïdes excrétés par la plante (Mulderet al, 2005). Si le mode de dialogue estidentique au cours des symbiosesactinorhiziennes alors il est nécessaired’activer les bactéries en les mettant aucontact de la plante pour qu’ellesproduisent le facteur symbiotique. Cesexpériences d’activation associées à destentatives de concentration du surnageantde Frankia par lyophilisation sont en coursdans notre laboratoire. L’obtention desurnageant activé et concentré permettrade répéter les tests d’induction des gènesCg12 et CgAUX1. Dans le cas où ces testss’avéreraient positifs, ils ouvriraient la voieà l’identification du signal symbiotique enutilisant ces gènes comme marqueurs.

51
CONCLUSION GÉNÉRALE – PERSPECTIVES
Nous avons étudié le rôle du transportde l’auxine au cours de la mise en place dela symbiose actinorhizienne chezCasuarina glauca. Deux homologues desgènes AUX-LAX impliqués dans letransport d’influx d’auxine ont étéidentifiés. L’analyse des séquences amontré une forte homologie de ces gènesavec AtAUX1 et AtLAX3 ce qui a permis deles nommer CgAUX1 et CgLAX3. Lesprofils d’expression de ces gènes sontégalement très conservés chezArabidopsis thaliana et C. glauca. Lasimilarité entre CgAUX1 et AtAUX1 a étéconfirmée par la capacité de CgAUX1 àcomplémenter le mutant aux1. L’analysedes profils d’expression a révélé uneassociation étroite entre l’expression deCgAUX1 et l’infection par Frankia ainsiqu’une différence d’expression deCgAUX1 entre le primordium de racine etde nodule. Afin d’identifier les gènesinduits en aval du transport d’influxd’auxine, nous avons participé àl’identification de ces cibles chez A.thaliana en analysant le mutant lax3. Cesétudes ont révélé que le remodelage de laparoi au cours de l’émergence était induitpar un signal auxinique sous dépendancedu gène AtLAX3. Par homologie, nousavons choisi comme candidat le gèneCg12 potentiellement impliqué dans leremodelage de la paroi et exprimé aucours de l’infection par la bactérie Frankia.L’expression de ce gène dans les cellulesinfectées semble dépendre en partie d’unsignal auxinique.
I. Le signal auxiniquepermet l’infection de lacellule végétale par desmicro-organismes
Nos résultats ont mis en évidence unlien entre le transport d’influx d’auxine etl’infection par la bactérie Frankia. Ces
résultats suggèrent un rôle important del’auxine dans la mise en place d’unprogramme symbiotique par la plante.Chez la plante actinorhizienne Elaeagnusumbellata, une protéine de type ARP estsurexprimée dans les nodules, enparticulier au niveau de la zone de fixationde l’azote (Kim et al, 2007). L’expressionde ce gène est induite par l’auxine ce quiconfirme la présence d’une signalisationauxinique dans ces cellules. En effet, unmutant de Bradyrhizobium japonicumsurproducteur d’auxine présente uneefficacité de nodulation plus élevée chezles Légumineuses (Kaneshiro et Kwolek,1985). Afin de confirmer cette hypothèse,il serait intéressant de déterminer sil’inhibition de la signalisation auxiniquepar surexpression d’une versiondominante négative d’une protéineAux/IAA (par exemple shy2/iaa3 ; Tian etal, 2002) perturbe l’infection.
L’auxine joue aussi un rôle importantdans la mise en place des symbiosesmycorhiziennes (Barker et Tagu, 2000). Eneffet, chez le maïs, les racinesendomycorhizées présentent des teneursimportantes en auxine (Jentschel et al,2006). De plus, un mutant surproducteurd’auxine d’Hebeloma cylindrosporumforme trois à cinq fois plusd’ectomycorhizes chez Pinus pinaster quele sauvage (Gay et al, 1994). Néanmoins,le rôle du transport d’influx d’auxine n’ajamais été étudié au cours de cessymbioses. La protéine CgAUX1 pourraitêtre impliquée dans la prise en charge del’auxine fongique au cours de l’infection.La plante tropicale C. glauca est capabled’intéragir avec différents champignonspour aboutir aux deux types de symbioses(Figure 50 ; Duponnois et al, 2003) ce quipermet une adaptation importante auxdifférents type de sols. Des analyses sonten cours dans notre laboratoire pourdéterminer le profil d’expression deCgAUX1 au cours de l’infection de C.

52
glauca par Pisolithus alba (ectomycorhizes)et Glomus intraradices (endomycorhizes).De plus, l’exctinction de l’expression deCgAUX1 par la technique d’ARNinterférent permettra de préciser le rôle dutransport d’influx d’auxine dans lessymbioses mycorhiziennes.
De façon plus générale, l’auxine estimpliquée dans les mécanismesd’infection par les bactéries pathogènes.En effet, de nombreuses bactéries du solproduisent des auxines (Lambrecht et al,2000). Parmi elles, certaines bactériesphytopathogènes utilisent cette auxinepour produire des tumeurs commePseudomonas savastanoi ou Erwiniaherbicola (Hutcheson et Kosuge, 1985 ;Manulis et al, 1998) ou au cours del’infection des tissus végétaux sans que lerôle précis de l’auxine ne soit connu.
Récemment, des études ont montré quel’activation de la signalisation auxiniqueaugmente la susceptibilité d’A. thaliana àl’infection par P. syringae (Navarro et al,2006). Les mécanismes de résistance de laplante passent par une diminution de laréponse auxinique. La perception d’unpeptide bactérien dérivé de la flagelline deP. syringae induit un micro-ARN quiréprime l’expression des récepteurs àl’auxine TIR1, AFB2 et AFB3 et permet delimiter l’infection par la bactérie. Il sembledonc que l’absence de perceptionauxinique constitue un verrou permettantà la cellule de bloquer l’infection. Unfacteur de transcription de type bZIPnommé BZI–1 est impliqué dans laréponse à l’auxine et se lie spécifiquementsur des éléments ACGT situés dans lepromoteur du gène GH3 induit parl’auxine (Heinekamp et al, 2004). Lors del’infection par P. syringae, l’expression deBZI-1 est augmentée ce qui suggère quel’activation de la signalisation auxiniqueouvre la voie à l’infection (Heinekamp et al,2002).
L’activation de la cellule par l’auxinesemble être un prérequis pour permettrel’infection que ce soit dans le cadre d’unesymbiose ou d’une infection pathogène.Cependant, l’auxine ne peut pas êtreconsidérée comme un signal spécifique.En effet, cette hormone intervient dans laplupart des mécanismesdéveloppementaux chez les plantes. Ilsemblerait plutôt que l’auxine constitueun signal d’activation des cellulesvégétales. La spécificité de l’activation estdéterminée par le type cellulaire, parexemple l’apport d’auxine à une cellule dupéricycle située en face d’un pôle dexylème aboutit à l’initiation duprimordium. Nous avons vu que dans lescellules du cortex et de l’épiderme, cetteactivation aboutit au remodelage de laparoi pour permettre l’émergence. Cemécanisme d’émergence estprobablement détourné par les bactériesphytopathogènes du sol afin de forcer leurpassage entre ces cellules. C’estcertainement la raison pour laquellel’induction d’AtLAX3 et des gènes deremodelage de la paroi dans l’épidermenécessite un signal plus fort que dans le
Figure 50. Processus de mise en place des symbiosesmycorhiziennesEctomycorhize. L’interaction débute par un échange designaux entre les deux partenaires (A). Les hyphess’accrochent à la surface racinaire et commencent à seramifier et à se renfler (B). L’ectomycorhize mature (C)présente trois structures typiques : le réseau de Hartig oùont lieu les échanges de nutriments (1), un manteau densequi couvre la racine (2) et un mycélium extraradiculaire quiexplore le sol (3).Endomycorhize. Stade asymbiotique : la germination de laspore est suivie d’une croissance limitée de l’hyphe (A).Stade présymbiotique : des signaux racinaires induisent lacroissance et la ramification de l’hyphe (B). Stadesymbiotique : colonisation intercellulaire et formation devésicules et arbuscules intracellulaires (C).La plante actinorhizienne Casuarina glauca peut réaliserces deux types de symbioses en intéragissant avecdifférents champignons du sol.

53
cortex. En effet, l’épiderme constitue lapremière barrière face aux agressions dela rhizosphère.
L’évolution des interactions pathogènesvers des interactions de type symbiotiquea pu conserver ce mécanisme d’infection.Un moyen pour la plante de limiter laprogression de la bactérie symbiotiquepourrait être de réguler sa sensibilité àl’auxine comme dans le cas desinteractions phytopathogènes. Lanécessité d’un signal symbiotiquespécifique d’une espèce est apparue aucours de évolution pour éviter auxbactéries non-symbiotiques de profiter dela situation. La présence d’un signalsymbiotique permet donc probablement àla cellule de l’épiderme ou du cortex des’engager vers un programmesymbiotique en réponse à l’activationauxinique. En l’absence de ce signalspécifique, l’activation par l’auxineengendrerait une réponse non-symbiotique telle que celle observée lorsde l’émergence de la racine latérale. Il estintéressant de constater que la bactérieFrankia n’infecte que les cellules del’épiderme puis du cortex (que ce soit dela racine ou plus tard du nodule) or ce sontces cellules qui chez Arabidopsis (et doncprobablement chez C. glauca si lemécanisme d’émergence est conservé)expriment des gènes de remodelage de laparoi en réponse à une activationauxinique. Ce mécanisme pourraitexpliquer pourquoi l’infection des cellulesdu péricycle n’est jamais observée, car cescellules ne sont pas programmées pourexprimer des gènes de remodelage de laparoi en réponse à l’auxine. Une autrehypothèse est que le signal symbiotiqueest perçu également dans ces cellulesmais bloque toute possibilité de réponseauxinique afin d’arrêter la progression dela bactérie vers les tissus vasculaires.
II. Le nodule actinorhizien :une racine latéralemodifiée ?
Le nodule actinorhizien est considérécomme une racine latérale modifiée(Pawlowski et Bisseling, 1996). En effet, leprimordium nodulaire est formé par
division des cellules du péricycle situéesen face du pôle de xylème et le nodulemature présente un méristème apical etun système vasculaire central. Néanmoins,nos résultats suggèrent qu’il existe desdifférences entre ces deux organes. Eneffet, le gène CgAUX1 est exprimé dans leprimordium de racine latérale mais estexclu du primordium nodulaire. Chez lesLégumineuses, il existe certainesressemblances entre le nodule et la racinelatérale. En effet, les gènes AUX/LAXprésentent des profils d’expressionsimilaires au cours de la formation de cesdeux organes (de Billy et al, 2001).Néanmoins, les cytokinines jouent un rôleopposé dans la formation de ces deuxorganes : inhibition de la formation desracines latérales et activation de laformation des nodules (Lohar et al, 2004 ;Eckardt, 2006 ; Gonzalez-Rizzo et al,2006). De plus, les nodules deLégumineuses qu’ils soient de typedéterminé ou indéterminé présentent unsystème vasculaire périphérique qui lesapparente d’avantage aux tiges qu’auxracines. Il est donc possible que différentsprogrammes de développement aient étérecrutés au cours de l’évolution pour lamise en place des symbiosesactinorhiziennes et Rhizobium-Légumineuses. Dans les deux cas,l’évolution de la symbiose s’estprobablement accompagnée d’une dérivedu programme génétique initial. Celaexplique pourquoi des différences existententre nodule actinorhizien et racinelatérale.
Le recrutement d’un programmegénétique de racine peut égalementpermettre la formation d’un nouvel organenon-symbiotique. Les racines protéoïdessont des touffes de racines latéralesmodifiées présentant une croissancelimitée. Elles sont caractéristiques desplantes de la famille des Protéacées mêmesi elles existent chez d’autres espèces(Neumann et Martinoia, 2002). Ellespermettent de mobiliser activement lesnutriments du sol comme le fer, lephosphate, le zinc et le manganèse enprovoquant des changements dans larhizosphère (pH, production d’exsudats etpotentiel redox). La capacité à former ces

54
racines protéoïdes n’est cependant paslimitée aux Protéacées, en effet dans leclade des Rosids cette propriété estretrouvée chez les Fagales (Betulacées,Casuarinacées et Myricacées), chez lesRosales (Elaeagnacées et Moracées) etchez les Fabales (Fabacées). La plantetropicale C. glauca est capable de formerces structures en cas de carence en fer eten phosphate. De façon surprenante, lesplantes actinorhiziennes ont toutes lacapacité de former ces racines protéoïdes(Lambers et al, 2006). Il semble doncexister un lien évolutif entre la capacité deformer ces racines et l’association avec labactérie Frankia. Cependant, aucune basemoléculaire de cette association n’a été
identifiée à ce jour. Il se pourrait que lacapacité à « dupliquer » le programmedévelopemental de racine latérale ait étéun prérequis pour la mise en place dunodule actinorhizien et des racinesprotéoïdes au cours de l’évolution.
Il serait intéressant de déterminer si letransport d’influx d’auxine est impliquédans la production des racines protéoïdesnotamment en analysant le profild’expression de CgAUX1 dans leursprimordia. En effet, la présence oul’absence d’expression de CgAUX1 dansces structures permettra de déterminer sileur programme de développement serapproche ou non de celui de la racinelatérale. L’extinction de l’expression de
CgAUX1 par ARN interférent permettraégalement de préciser l’implication de ce
gène dans la mise en place de la racineprotéoïde.

55
MATÉRIEL ET MÉTHODES
I. Matériel1) Matériel végétalLes graines de Casuarina glauca
proviennent des Etats-Unis (Réf. 06003,Société Carter Seeds, Californie, USA). Lesgraines d’Allocasuarina verticillata sontd’origine australienne (Société Versepuy,Le Puy-en-Velay, France).
Les graines d’Arabidopsis thalianaécotype Columbia-0 (Col0), écotypeLandsberg erecta (Ler) et mutants (aux1-22, lax3, aux1lax3 et shy2-2) ont étéfournies par le NASC (NottinghamArabidopsis Stock Center).
2) Matériel bactérienLa souche de Frankia utilisée pour
inoculer C. glauca et A. verticillata est lasouche CcI3 qui est maintenue aulaboratoire.
Les souches d’Escherichia coli utiliséessont soit JM109 (Promega) dont legénotype est : e14–(McrA–) recA1 endA1gyrA96 thi-1 hsdR17 (rK
– mK+) supE44
relA1 Δ(lac-proAB) [F´ traD36 proABlacIqZΔM15], soit XL1-blue dont legénotype est recA1 endA1 gyrA96 thi-1hsdR17 supE44 relA1 lac [F´ proABlacIqZΔM15 Tn10 (Tetr)].
Les souches d’Agrobacteriumtumefaciens utilisées sont de génotypeC58C1. Elles portent le plasmide désarmépGV3101 pour la transformationgénétique d’A. thaliana ou pGV2260 pourla transformation génétique de C. glaucaet A. verticillata. Elles sont résistantes à larifampicine (10 mg.L-1).
3) Matériel nucléique
a) PlasmidesLes stratégies de clonage utilisées pour
les constructions principales sont donnéesdans les figures 51 et 52.
Le plasmide binaire utilisé pour réaliserles fusions promoteur-GUS est pBI101.3(Clontech).
Le plasmide binaire utilisé pour réaliserles fusions promoteur-GFP estpBI101.mGFP5ER dérivé du pBI101.3(Svistoonoff et al, 2003).
Le plasmide binaire utilisé pour réaliserles constructions pour la complémentationfonctionnelle de mutants d’A. thaliana estpMOG402 ou son dérivé pMOG AUX1ORFL contenant le promoteur (1,6kb) et leterminateur (0,3 kb) d’AtAUX1.
Ces trois vecteurs binaires portent ungène de résistance à la kanamycine (50mg.L-1) à expression procaryotique et ungène de résistance à la kanamycine (NptII)sous dépendance du promoteur viral 35Sà expression eucaryotique.
Les clonages en routine de produit PCRsont réalisés à l’aide de pGEM-T easy(Promega) qui confère la résistance àl’ampicilline (100 mg.L-1) ou de pCR2.1-TOPO (Invitrogen) qui confère la résistanceà l’ampicilline (100 mg.L-1) et à lakanamycine (50 mg.L-1).
b) Amorces pour la PCRLes séquences des amorces utilisées
sont rassemblées dans le tableau I. Lesséquences sont données de 5’ vers 3’. Lesamorces ont été dessinées à l’aide dulogiciel Beacon Designer (Premier BiosoftInternational) et synthétisées parEurogentec.
II. Méthodes1) Milieux de culture (plantes)
a) Casuarina glaucaLes graines de C. glauca sont mises à
germer en serres sur un mélangeterreau/vermiculite (1/1 , v/v). Un moisenviron après germination, les plantulessont transférées pour culture

56
hydroponique sur solution nutritive BD(Broughton et Dilworth, 1971) avec ajoutd’azote (KNO3 5 mM). Trois semainesaprès transfert, les plantes sont sevrées enazote pendant une semaine puis sontinoculées par Frankia toujours sur milieusans azote.
b) Arabidopsis thalianaLes graines d’A. thaliana sont mises à
germer en serres ou en salle de culture(60% d’humidité, cycles 16h jour/8h nuit,intensité lumineuse 45 µE) sur mélangeterreau/vermiculite (1/1 , v/v)éventuellement additionné d’engrais àdiffusion lente. Pour la culture stérile, les
graines sont traitées avec du Domestos20% pendant 10 minutes puis rincées troisfois avec de l’eau stérile. Elles sontdéposées sur boîtes horizontales avecmilieu 1/2MS (Murashige et Skoog, 1962)additionné des vitamines B5 de Gamborg(Duchefa), plant agar 0,8% (m/v ; Kalys)ajusté à pH 5,6 ou bien sur boîtesverticales avec milieu 1/2MS additionnédes vitamines B5 de Gamborg, agar HP1,2% (m/v ; Kalys) ajusté à pH 5,6. Lesboîtes sont placées à 4°C pendant deuxjours pour synchroniser la germination etensuite transférées en salle de culture(60% d’humidité, cycles 16h jour/8h nuit,intensité lumineuse 45 µE).
Figure 51. Construction desvecteurs binaires ProCgAUX1:GUS,ProCgLAX3:GUS et ProAtAUX1:CgAUX1
57
2) Milieux de culture (bactéries)Les souches d’E. coli et d’A.
tumefaciens sont cultivées sur milieu LBliquide ou solide (agar 1,5% m/v) à 37°C et28°C respectivement (composition dumilieu LB en tableau II). Elles sontconservées à –80°C dans une solution deglycérol 15%.
La bactérie Frankia souche CcI3 est
cultivée sur milieu BAP-PCM à 28°C et àl’obscurité sous agitation (composition dumilieu BAP-PCM en tableau III). Aumoment du repiquage, les bactéries sont
Figure 52. Construction desvecteurs binairesProAtAUX1:CgLAX3 etProAtLAX3:CgLAX3

58
centrifugées 15 minutes à 8000g puisreprises dans de l’eau stérile et les hyphessont brisés par une dizaine de passagesdans une seringue munie d’une aiguillestérile Terumo (0,7 x 30mm). La solutioncontenant les bactéries est utilisée pourensemencer le nouveau milieu de cultureou pour inoculer les plantes une semaineaprès sevrage.
3) Transformation génétique desplantes
a) Casuarina glauca et Allocasuarinaverticillata
Les graines de C. glauca et A. verticillatasont scarifiées par trempage dans H2SO496% pendant 2 minutes, rincées à l’eaucourante 30 minutes, rincées dans duTween 80 (quelques gouttes dans environ50mL H20), rincées rapidement dans de
l’éthanol 75%, désinfectées à l’hypochloritede calcium 5% pendant 35 minutes,rincées à l’eau stérile deux fois puislaissées dans l’eau stérile toute la nuit. Lesgraines sont ensuite mises à germer surmilieu H (Hoagland et Arnon, 1938) diluéau 1/4. Les fragments d’épicotyles sontplacés en contact des agrobactéries surmilieu MSC liquide (la composition desdifférents milieux MSC est présentée dansle tableau IV) puis la coculture est réaliséependant trois jours sur milieu MSC solide.Les explants sont rincés trois fois avec duMSC liquide pendant une heure puisplacés sur MSC avec régulateurs etantibiotiques : céfotaxime (250 mg.L-1) etkanamycine (25 mg.L-1 pour C. glauca et50 mg.L-1 pour A. verticillata). Aprèsquinze jours, les concentrations enkanamycine sont doublées et le milieu estrenouvelé toutes les trois semaines. Les
Tableau I. Séquences des amorcesutilisées pour la PCRLes séquences sont données dans lesens 5’ vers 3’.

59
cals apparaissent après un à deux mois. Lacéfotaxime est progressivement retirée aubout de six mois. Lorsque les calsatteignent 5 mm environ, ils sonttransférés en tube en verre. Les rameauxde C. glauca sont enracinés par chocauxinique sur milieu MSC-IBA pendantdeux jours puis transférés sur milieu MSCavec charbon actif. Les rameaux d’A.verticillata sont enracinés par chocauxinique sur milieu MSC-NAA pendantdeux jours puis transférés sur milieu MSC-enracinement liquide. Dans les deux cas,les premières racines apparaissent au boutd’environ une semaine. Pour l’inoculationpar Frankia, les plants de C. glauca sonttransférés en hydroponie non stérile (voirMilieux de culture) et traités comme desplantes non transformées. Les plants d’A.verticillata sont transférés en tube en verrecontenant une gélose inclinée (milieu MSCpour tube) et du milieu H liquide avecazote. Le sevrage en azote est effectuéune semaine avant inoculation par Frankia.
La multiplication végétative desrameaux de C. glauca est réalisée parbouturage. Les rameaux (environ 3 à 5cm) sont placés 24 heures dans unesolution de NAA 0,1 g.L-1 puis transférésdirectement en pot pour culturehydroponique sur milieu BD. Lespremières racines apparaissent au boutd’une semaine environ.
b) Arabidopsis thalianaLa transformation génétique d’A.
thaliana est réalisée par la techniqued’imprégnation des boutons floraux(Clough et Bent, 1998). Une premièreculture de bactérie (LB 5 mL, kanamycine50 mg.L-1, rifampicine 10 mg.L-1) estlancée à partir de la solution stockpendant 16 heures à 28°C sous agitation.Une deuxième culture (LB 500 mLkanamycine 50 mg.L-1) est inoculée avec500 µL de la première, elle est laisséependant 16 heures à 28°C sous agitation.Les bactéries sont culotées parcentrifugation (6000 g, 15 minutes àtempérature ambiante) et reprises dansune solution de saccharose 5% (m/v) etsilwet L-77 0,05% (v/v). Les boutonsfloraux (plantes âgées d’un mois environ)
sont trempés dans la solution pendant 5secondes environ. Les plantes sont placéesà l’obscurité sous mini-serres pendant 16heures puis replacées dans le phytotron.Deux imprégnations sont réalisées à septjours d’intervalle afin d’augmenterl’efficacité de la transformation.
4) Transformation génétique desbactéries
a) Escherichia coliLes bactéries XL1-blue sont rendues
compétentes à la transformation par chocthermique (Hanahan, 1983). Une culturede 100 mL de bactéries (densité optique à600 nm entre 0,3 et 0,4) est centrifugée10 minutes à 4000 g à 4°C. Le culot estrepris dans 30 mL de la première solution(RbCl 100 mM, MnCl2 50 mM, Acétate depotassium 30 mM, CaCl2 10 mM, glycérol15 %, pH 5,8). Après centrifugation 10minutes à 4000 g à 4°C, le culot est reprisdans 7 mL de la deuxième solution(MOPS 10 mM, RbCl 10 mM, CaCl2 75 mM,glycérol 15 %, pH 6,8). Les bactérieslaissées 15 minutes sur glace sontaliquotées par 150 µL dans des tubes de1,5 mL et immédiatement plongées dansl’azote liquide (-180°C). Les bactériescompétentes sont conservées à –80°Cjusqu’à utilisation. Les bactériescommerciales JM109 (Promega) sontprêtes à l’emploi.
Les plasmides sont intégrés dans lesbactéries compétentes par chocthermique. Un volume de 50 µL debactéries est mélangé avec environ 10 µLde plasmide et laissé sur glace une minutepuis à 42°C pendant 50 secondes puisretour sur glace deux minutes. Un volumede 950 µL de LB est ajouté et les bactériessont placées à 37°C sous agitationpendant une heure. Les bactériestransformées sont sélectionnées sur boîtesde LB agar contenant l’antibiotiqueapproprié.
b) Agrobacterium tumefaciensLes agrobactéries sont rendues
compétentes à l’électroporation. Uneculture d’un litre de bactéries (densitéoptique à 600 nm entre 0,6 et 0,9) est

60
laissée sur glace pendant 15 minutes puiscentrifugée 10 minutes à 3000 g à 4°C. Leculot est repris successivement dans unlitre d’eau stérile froide, 500 mL d’eaustérile froide, deux fois dans 20 mL deglycérol 10% froid après centrifugationssuccessives de 10 minutes à 3000 g à 4°C.Les bactéries sont aliquotées par 50 µLdans des tubes de 1,5 mL etimmédiatement plongées dans l’azoteliquide (-180°C). Les bactériescompétentes sont conservées à –80°Cjusqu’à utilisation.
Les bactéries compétentes sonttransformées par électroporation. Unvolume de 50 µL de bactéries est mélangéavec environ 10 µL de plasmide dilué etplacé dans la cuve de l’électroporateur. Lesbactéries sont soumises à un chocélectrique de 2,5 kV, 192 W et 40 µF puisun volume de 950 µL de LB est ajouté etles bactéries sont placées à 28°C sousagitation pendant deux heures. Lesbactéries transformées sont sélectionnéessur boîtes de LB agar contenantl’antibiotique approprié.
5) Biologie moléculaire
a) Extraction des acides nucléiquesLes ARN totaux de C. glauca sont
extraits selon la méthoded’ultracentrifugation sur coussin dechlorure de césium (Chirgwin et al, 1979).Les ARN totaux d’A. thaliana sont extraitsà l’aide du kit RNeasy (Qiagen) selon lesinstructions du fabricant.
L’ADN génomique de C. glauca estextrait selon la méthode au MATAB (Ky etal, 2000). L’ADN génomique d’A. thalianaest extrait à l’aide du kit Dneasy (Qiagen).
L’ADN plasmidique est extrait descultures bactériennes à l’aide du kitMiniPrep (Qiagen).
b) Traitement DNAsePour utilisation en RT-PCR, les ARN
sont soumis à un traitement DNAse àl’aide du kit DNA-free (Ambion) afin desupprimer toute trace d’ADN génomique.
c) Purification des ARN polyALes ARN poly-adénylés sont purifiés à
l’aide du kit mRNA purification (AmershamBiosciences) à partir des ARN totaux(environ 40 µg).
d) Northern blotLes ARN totaux sont soumis à un
Northern blot à l’aide du kit NorthernMax-Gly (Ambion). Après électrophorèse, lesARN sont transférés sur membrane ennylon (solution de transfert SSC 10X) parcapillarité.
e) Southern blotL’ADN génomique de C. glauca (environ
15 µg) est digéré par des enzymes derestriction (15 unités.µg-1 ADNg à 37°Ctoute la nuit). L’ADN digéré est placé sousélectrophorèse (1V.cm-1) dans un geld’agarose 1% (m/v dans tampon TBE 1X)dans du tampon TBE 1X. Aprèsélectrophorèse, l’ADN est visualisé parcoloration au bromure d’éthidium etéclairage aux rayons ultraviolets. Le gel esttraité pour dénaturer l’ADN (NaCl 1,5 M ;NaOH 0,5 M) pendant une heure sousagitation légère à température ambiante.Puis le gel est placé dans une solution deneutralisation (NaCl 1,5 M ; Tris-HCl 1MpH 7,4) pendant une heure. L’ADN estensuite transféré sur une membrane ennylon (solution de transfert SSC 10X) parcapillarité.
f) Hybridation des membranesLa membrane de Northern ou Southern
blot est séchée et les acides nucléiquessont fixés par exposition aux rayonsultraviolets (120 mJ.cm-2). La membraneest traitée en pré-hybridation au minimum3 heures à 65°C (Denhardt’s 5X, SSC 6X,SDS 0,5%, ADN de sperme de harengdénaturé 0,1 mg.mL-1). La sonde (environ25 ng de produit PCR purifié) estradiomarquée au α32P-dCTP (50 µCi) par latechnique de l’amorçage aléatoire(random priming) à l’aide du kitMegaprime™ DNA labelling systems(Amersham Biosciences). La sonde estajoutée pour hybridation toute la nuit. Lasolution d’hybridation est éliminée et lamembrane est lavée à l’aide de solutions

61
de lavage de stringence croissante (SSC2X, SDS 0,1% ; SSC 1X, SDS 0,1% ; SSC0,5X, SDS 0,1%). La membrane est alorsplacée au contact d’un filmautoradiographique (Kodak) pendant unenuit dans une cassette à –80°C. Le film estrévélé (solution de révélation, Kodak) puisfixé (solution de fixation, Kodak).
g) Transcription inverseLes ARN totaux (250 ng à 1µg) sont
rétro-transcrits à l’aide du kit ReverseTranscription System (Promega) ou du kitSuperscript III Reverse Transcriptase(Invitrogen) selon les instructions dufabricant.
h) RACE-PCR Les ADNc correspondant aux fragments
de gènes clonés par PCR sont obtenus paramplification rapide des extrémités del’ADNc (RACE-PCR) à l’aide du kitMarathon™ cDNA amplification (Clontech)selon les instructions du fabricant.
i) Marche sur l’ADN génomiqueLes séquences promotrices des gènes
sont obtenues par marche sur l’ADNgénomique à l’aide du kit UniversalGenomeWalker (Clontech).
j) RT-PCR quantitativeTrois réactions indépendantes de
transcription inverse sont effectuées àpartir de 200 à 400 ng d’ARN totaux. Laréaction de RT-PCR quantitative estréalisée sur un appareil Mx3005P(Stratagene) en utilisant le mélangeFullVelocity SYBR Green QPCR Master Mix(Stratagene). Pour l’amplification, leséchantillons sont chauffés 5 minutes à95°C, puis soumis à 40 cycles dedénaturation (95°C pendant 10 secondes)et hybridation-extension (60°C pendant30 secondes). Une courbe de dissociationest réalisée en chauffant les échantillonsde 60 à 95°C afin de déterminer la Tm desproduits amplifiés.
k) Séquençage de l’ADNLes séquençages d’ADN sont réalisés
par la société MWG à partir de 1µg d’ADNplasmidique.
l) Analyse des séquencesLes séquences sont soumises au BLAST
(Altschul et al, 1990) pour comparaison.Les arbres phylogénétiques sont dessinésavec le logiciel Treecon v1.3b. Lesalignements sont réalisés avec leprogramme ClustalW.
6) Histologie
a) Coloration GUSLes tissus prélevés ou les coupes
épaisses réalisées au vibratome sontplongés dans une solution de X-Glc (X-Glc0,5g.L-1, diméthylformamide 0,5% v/v,EDTA 1mM, Triton X-100 0,5% v/v,Na2PO4 500mM pH 7). Pour Arabidopsis,l’ajout de K3Fe(CN)6 et K4Fe(CN)6 0,5 mMpermet de limiter la diffusion des cristauxbleus. L’incubation est réalisée pendant 2à 16 heures à 37°C suivant l’expression dupromoteur.
b) FixationLes tissus sont fixés dans de l’éthanol
70% (plusieurs bains pendant au moins unjour). Ils sont ensuite traités pour inclusionen résine ou progressivement transférésdans du glycérol 50% pour conservation(éthanol 50% glycérol 10% v/v pendant 2heures, éthanol 30% glycérol 30% pendant2 heures puis glycérol 50% pendant 2heures).
c) Inclusion en résineLes tissus sont inclus dans de l’agarose
avant inclusion en résine afin de faciliterleur orientation. Les tissus sontprogressivement amenés dans de l’eau(bains successifs de 15 minutes dans del’éthanol 50%, 30% puis 10%). Les tissussont ensuite inclus dans de l’agarose 1,2%(m/v) tiède et les blocs d’agarose sontfaçonnés au scalpel.
Les blocs d’agarose sont déshydratéspar bains successifs de 15 minutes dansde l’éthanol 10%, 30%, 50%, 75%, 95% puistrois fois 100%. Les échantillonsdéshydratés sont inclus dans une résineTechnovit 7100 (Heraeus Kulzer,Wehrheim, Allemagne) selon instructionsdu fabricant.

62
d) Coupes épaissesLes coupes épaisses (40 à 60 µm) sont
réalisées sur des tissus frais inclus dans del’agarose 3% (m/v) à l’aide d’un vibratomeVT1000E (Leica).
e) Coupes finesLes coupes fines (3 à 6 µm) sont
réalisées sur bloc de résine à l’aide d’unmicrotome MT355S (Leica).
f) Montage et observationLes tissus non coupés (racine
d’Arabidopsis) sont directement montésentre lame et lamelle dans une solution deglycérol 50%. Les coupes fines sontmontées sur lame dans une solution deClearium mountant (Surgipath) et lescoupes épaisses sont montées dans del’eau. Les lames sont observées avec unmicroscope Leitz DMRB. Les images sontprises avec une caméra numériqueDFC300FX (Leica). Le traitement desimages est réalisé avec le logicielGraphicConverter (lemkesoft.com).
7) Analyses phénotypiques
a) GravitropismeLe phénotype gravitropique des plants
d’Arabidopsis est testé sur boîte verticale.Les plantes sont mises à germer sur boitespendant une semaine puis les boites sonttournées à 90°C, l’angle de réorientationdes apex racinaires est estimévisuellement 24 heures après stimulusgravitropique.
b) Résistance au 2,4DLa résistance au 2,4D est testée sur
boîte verticale. Les plantes sont mises à
germer sur milieu avec ou sans auxine etla longueur de la racine primaire estmesurée 5 jours près germination avecl’aide du logiciel ImageJ 1.37v. Leslongueurs de racines sont expriméesrelativement à la longueur des racinestémoin (milieu sans 2,4D).
8) Dosages
a) Dosage des ARNPour un dosage approximatif, l’ADN et
l’ARN sont dosés par spectrophotométrieà 260 nm. Pour un dosage plus précis, lesARN sont dosés avec du RibogreenReagent (Stratagene) à l’aide de l’appareilMx3005P (Stratagene) par comparaisonavec une gamme étalon d’ARN.
b) Dosage des protéines totalesLes protéines totales de Frankia sont
dosées par la coloration de Bradford(Bradford, 1976) sur milieu de culturesoumis à une sonication rapide (troisimpulsions à 50% d’intensité). La densitéoptique à 595 nm (vérifier) est mesurée 5minutes après mélange avec le réactif deBradford (1/1, v/v). La quantité deprotéines est déterminée par comparaisonavec une gamme étalon de sérumalbumine bovine.
c) Dosage des auxinesLa quantité d’auxine (IAA et PAA) dans
les racines et nodules de C. glauca et dansle surnageant de culture de Frankia estdéterminée par HPLC couplée à laspectrométrie de masse (Kowalczyk etSandberg, 2001). Les dosages sonteffectués par le D. Helena Fernandez(Université d’Oviedo, Espagne).

63
RÉFÉRENCES BIBLIOGRAPHIQUES
Altschul SF, Gish W, Miller W, Myers EW et Lipman DJ(1990) Basic local alignment search tool. J Mol Biol.215: 403-410.
Ané JM, Kiss GB, Riely BK, Penmetsa RV, Oldroyd GE, AyaxC, Lévy J, Debellé F, Baek JM, Kalo P, Rosenberg C,Roe BA, Long SR, Dénarié J et Cook DR (2004)Medicago truncatula DMI1 required for bacterial andfungal symbioses in legumes. Science. 303: 1364-1367.
Angulo Carmona AF (1974) La formation des nodulesfixateurs d'azote chez Alnus glutinosa (L.). Acta BotNeerl. 23: 257-303.
Arabidopsis Genome Initiative (2000) Analysis of thegenome sequence of the flowering plant Arabidopsisthaliana. Nature. 408: 796-815.
Arp DJ (2000) The nitrogen cycle. In Prokaryotic nitrogenfixation : a model system for the analysis of abiological process. Triplett, EW (ed) Horizon ScientificPress, Wymondham, UK.
Badescu GO et Napier RM (2006) Receptors for auxin: will itall end in TIRs? Trends Plant Sci. 11: 217-223.
Baier R, Schiene K, Kohring B, Flaschel E et Niehaus K(1999) Alfalfa and tobacco cells react differently tochitin oligosaccharides and sinorhizobium melilotinodulation factors. Planta. 210: 157-164.
Barker DG, Bianchi S, Blondon F, Dattée Y, Duc G, Essad P,Flament P, Galuxy P, Génier G, Muel X, Toureur J,Dénarié J et Huguet T (1990) Medicago truncatula,a model plant for studying the molecular genetics ofRhizobium-legume symbiosis. Plant Mol Biol Rep. 8:40-49.
Barker SJ et Tagu D (2000) The Roles of Auxins andCytokinins in Mycorrhizal Symbioses. JOURNAL OFPLANT GROWTH REGULATION. 19: 144-154.
Bartel B, LeClere S, Magidin M et Zolman BK (2001) Inputsto the active indole-3-acetic acid pool : de novosynthesis, conjugate hydrolysis, and indole-3-butyric acid beta-oxidation. Journal Plant GrowthRegulation. 20: 198-216.
Beeckman T, Burssens S et Inzé D (2001) The peri-cell-cycle in Arabidopsis. J Exp Bot. 52: 403-411.
Beemster GT, Fiorani F et Inzé D (2003) Cell cycle: the keyto plant growth control? Trends Plant Sci. 8: 154-158.
Benfey PN et Scheres B (2000) Root development. Curr Biol.10: R813-R815.
Benková E, Michniewicz M, Sauer M, Teichmann T,Seifertová D, Jürgens G et Friml J (2003) Local,efflux-dependent auxin gradients as a commonmodule for plant organ formation. Cell. 115: 591-602.
Bennett MJ, Marchant A, Green HG, May ST, Ward SP,Millner PA, Walker AR, Schulz B et Feldmann KA(1996) Arabidopsis AUX1 gene: a permease-likeregulator of root gravitropism. Science. 273: 948-950.
Benoit LF et Berry AM (1997) Flavonoid-like compoundsfrom red alder (Alnus rubra) influence nodulation by
Frankia (Actinomycetales). Physiol Plant. 99: 588-593.
Benson DR et Silvester WB (1993) Biology of Frankiastrains, actinomycete symbionts of actinorhizalplants. Microbiol Rev. 57: 293-319.
Benson DR, et Clawson ML (2000) Evolution of theactinorhizal plant symbiosis. In Prokaryotic nitrogenfixation : a model system for the analysis of abiological process. Triplett, EW (ed) Horizon ScientificPress, Wymondham, UK.
Berg RH (1999a) Frankia forms infection threads. Can J Bot.77: 1327-1333.
Berg RH (1999b) Cytoplasmic bridge formation in thenodule apex of actinorhizal root nodules. Can J Bot.77: 1351-1357.
Berry AM, et Sunnel LA (1990) The infection process andnodule development. In The biology of Frankia andActinorhizal plants. pp 61-81. Schwintzer, CR andTjepkema JD (ed) Academic Press, New York.
Berry AM, McIntyre L et McCully M (1986) Fine structure ofroot hair infection leading to nodulation in theFrankia-Alnus symbiosis. Can J Bot. 64: 292-305.
Bhalerao RP, Eklöf J, Ljung K, Marchant A, Bennett MJ etSandberg G (2002) Shoot-derived auxin is essentialfor early lateral root emergence in Arabidopsisseedlings. Plant J. 29: 325-332.
Blakeslee JJ, Peer WA et Murphy AS (2005) Auxintransport. Curr Opin Plant Biol. 8: 494-500.
Bohlool BB, Ladha JK, Garrity DP et George T (1992)Biological nitrogen fixation for sustainableagriculture. A perspective. Plant Soil. 141: 1-11.
Boot KJM, van Brussel AAN, Tak T, Spaink HP et Kijne JW(1999) Lipochitin oligosaccharides from Rhizobiumleguminosarum bv. viciae reduce auxin transportcapacity in Vicia sativa subsp. nigra. Mol PlantMicrobe Interact. 12: 839-844.
Boutté Y, Crosnier MT, Carraro N, Traas J et Satiat-Jeunemaitre B (2006) The plasma membranerecycling pathway and cell polarity in plants: studieson PIN proteins. J Cell Sci. 119: 1255-1265.
Bradford MM (1976) A rapid and sensitive method for thequantitation of microgram quantities of proteinutilizing the principle of protein-dye binding. AnalBiochem. 72: 248-254.
Brewin NJ (2004) Plant cell wall remodelling in theRhizobium-Legume symbiosis. Critical Reviews inPlant Sciences. 23: 293-316.
Broughton WJ et Dilworth MJ (1971) Control ofleghaemoglobin synthesis in snake beans. BiochemJ. 125: 1075-1080.
Brown DE, Rashotte AM, Murphy AS, Normanly J, TagueBW, Peer WA, Taiz L et Muday GK (2001)Flavonoids act as negative regulators of auxintransport in vivo in arabidopsis. Plant Physiol. 126:524-535.
Callaham D et Torrey JG (1977) Prenodule formation andprimary nodule development in roots of Comptonia(Myricaceae). Can J Bot. 51: 2306-2318.

64
Callaham D, Newcomb W, Torrey JG et Peterson RL (1979)Root hair infetion in actinomycete-induced rootnodule initiation in Casuarina, Myrica andComptonia. Bot Gaz. 140: S1-S9.
Cannon SB, Sterck L, Rombauts S, Sato S, Cheung F,Gouzy J, Wang X, Mudge J, Vasdewani J, Scheix T,Spannagl M, Monaghan E, Nicholson C,Humphray SJ, Schoof H, Mayer KF, Rogers J,Quétier F, Oldroyd GE, Debellé F, Cook DR, RetzelEF, Roe BA, Town CD, Tabata S, Van de Peer Y etYoung ND (2006) Legume genome evolutionviewed through the Medicago truncatula and Lotusjaponicus genomes. Proc Natl Acad Sci U S A. 103:14959-14964.
Capoen W, Goormachtig S, De Rycke R, Schroeyers K etHolsters M (2005) SrSymRK, a plant receptoressential for symbiosome formation. Proc Natl AcadSci U S A. 102: 10369-10374.
Casimiro I, Beeckman T, Graham N, Bhalerao R, Zhang H,Casero P, Sandberg G et Bennett MJ (2003)Dissecting Arabidopsis lateral root development.Trends Plant Sci. 8: 165-171.
Casimiro I, Marchant A, Bhalerao RP, Beeckman T,Dhooge S, Swarup R, Graham N, Inzé D, SandbergG, Casero PJ et Bennett M (2001) Auxin transportpromotes Arabidopsis lateral root initiation. PlantCell. 13: 843-852.
Cassab GI (1998) PLANT CELL WALL PROTEINS. Annu RevPlant Physiol Plant Mol Biol. 49: 281-309.
Cérémonie H, Cournoyer B, Maillet F, Normand P etFernandez MP (1998) Genetic complementation ofrhizobial nod mutants with Frankia DNA: artifact orreality? Mol Gen Genet. 260: 115-119.
Cérémonie H, Debellé F et Fernandez MP (1999) Structuraland functional comparison of Frankia root hairdeforming factor and rhizobia Nod factor. Can J Bot.77: 1293-1301.
Chirgwin JM, Przybyla AE, MacDonald RJ et Rutter WJ(1979) Isolation of biologically active ribonucleic acidfrom sources enriched in ribonuclease. Biochem. 18:5294-5299.
Clough SJ et Bent AF (1998) Floral dip: a simplified methodfor Agrobacterium-mediated transformation ofArabidopsis thaliana. Plant J. 16: 735-743.
Cohen JD, Slovin JP et Hendrickson AM (2003) Twogenetically discrete pathways convert tryptophan toauxin: more redundancy in auxin biosynthesis.Trends Plant Sci. 8: 197-199.
Coronado C, Zuanazzi J, Sallaud C, Quirion JC, Esnault R,Husson HP, Kondorosi A et Ratet P (1995) AlfalfaRoot Flavonoid Production Is Nitrogen Regulated.Plant Physiol. 108: 533-542.
Cosgrove DJ (2000) Loosening of plant cell walls byexpansins. Nature. 407: 321-326.
Cullimore JV, Ranjeva R et Bono JJ (2001) Perception oflipo-chitooligosaccharidic Nod factors in legumes.Trends Plant Sci. 6: 24-30.
D'Agostino IB, Deruère J et Kieber JJ (2000)Characterization of the response of the Arabidopsisresponse regulator gene family to cytokinin. PlantPhysiol. 124: 1706-1717.
Dawson JO (1990) Interactions among actinorhizal andassociated plant species. In The biology of Frankia
and actinorhizal plants. pp 299-316. Schwintzer, CRand Tjepkema JD (ed) Academic Press, New York.
de Billy F, Grosjean C, May S, Bennett M et Cullimore JV(2001) Expression studies on AUX1-like genes inMedicago truncatula suggest that auxin is requiredat two steps in early nodule development. Mol PlantMicrobe Interact. 14: 267-277.
de Faria SM, Lewis GP, Sprent JI et Sutherland JM (1989)Occurence of nodulation in the leguminosae. NewPhytol. 111: 607-619.
De Smet I, Tetsumura T, De Rybel B, Frei Dit Frey N,Laplaze L, Casimiro I, Swarup R, Naudts M,Vanneste S, Audenaert D, Inzé D, Bennett MJ etBeeckman T (2007) Auxin-dependent regulation oflateral root positioning in the basal meristem ofArabidopsis. Development. 134: 681-690.
De Smet I, Vanneste S, Inzé D et Beeckman T (2006)Lateral root initiation or the birth of a new meristem.Plant Mol Biol. 60: 871-887.
Dharmasiri N, Dharmasiri S et Estelle M (2005a) The F-box protein TIR1 is an auxin receptor. Nature. 435:441-445.
Dharmasiri N, Dharmasiri S, Weijers D, Lechner E,Yamada M, Hobbie L, Ehrismann JS, Jürgens G etEstelle M (2005b) Plant development is regulated bya family of auxin receptor F box proteins. Dev Cell.9: 109-119.
Dharmasiri S, Swarup R, Mockaitis K, Dharmasiri N, SinghSK, Kowalchyk M, Marchant A, Mills S, SandbergG, Bennett MJ et Estelle M (2006) AXR4 Is Requiredfor Localization of the Auxin Influx Facilitator AUX1.Science. 312: 1218-1220.
Dolan L et Scheres B (1998) Root pattern: shooting in thedark? Semin Cell Dev Biol. 9: 201-206.
Dolan L, Janmaat K, Willemsen V, Linstead P, Poethig S,Roberts K et Scheres B (1993) Cellular organisationof the Arabidopsis thaliana root. Development. 119:71-84.
Dommergues Y, Duhoux E et Diem HG (1999) Les arbresfixateurs d'azote. Espace 34 (ed) Montpellier.
Dubrovsky JG, Doerner PW, Colón-Carmona A et Rost TL(2000) Pericycle cell proliferation and lateral rootinitiation in Arabidopsis. Plant Physiol. 124: 1648-1657.
Dubrovsky JG, Rost TL, Colón-Carmona A et Doerner P(2001) Early primordium morphogenesis duringlateral root initiation in Arabidopsis thaliana. Planta.214: 30-36.
Duponnois R, Diédhiou S, Chotte JL et Sy MO (2003)Relative importance of the endomycorrhizal and (or)ectomycorrhizal associations in Allocasuarina andCasuarina genera. Can J Microbiol. 49: 281-287.
Eckardt NA (2006) Medicago truncatula CRE1 CytokininReceptor Regulates Nodulation and Lateral RootDevelopment. Plant Cell. 18: 2419.
Endre G, Kereszt A, Kevei Z, Mihacea S, Kaló P et Kiss GB(2002) A receptor kinase gene regulating symbioticnodule development. Nature. 417: 962-966.
Fang Y et Hirsch AM (1998) Studying early nodulin geneENOD40 expression and induction by nodulationfactor and cytokinin in transgenic alfalfa. PlantPhysiol. 116: 53-68.

65
Foucher F et Kondorosi E (2000) Cell cycle regulation in thecourse of nodule organogenesis in Medicago. PlantMol Biol. 43: 773-786.
Franche C, Laplaze L, Duhoux E et Bogusz D (1998)Actinorhizal symbioses : recent advances in plantmolecular and genetic transformation studies. Crit.Rev. Plant Sci.. 17: 1-28.
Friml J (2003) Auxin transport - shaping the plant. CurrOpin Plant Biol. 6: 7-12.
Friml J et Palme K (2002) Polar auxin transport-oldquestions and new concepts? Plant Mol Biol. 49:273-284.
Fukaki H, Nakao Y, Okushima Y, Theologis A et Tasaka M(2005) Tissue-specific expression of stabilizedSOLITARY-ROOT/IAA14 alters lateral rootdevelopment in Arabidopsis. Plant J. 44: 382-395.
Fukaki H, Tameda S, Masuda H et Tasaka M (2002) Lateralroot formation is blocked by a gain-of-functionmutation in the SOLITARY-ROOT/IAA14 gene ofArabidopsis. Plant J. 29: 153-168.
Gage DJ (2004) Infection and invasion of roots by symbiotic,nitrogen-fixing rhizobia during nodulation oftemperate legumes. Microbiol Mol Biol Rev. 68: 280-300.
Gage DJ et Margolin W (2000) Hanging by a thread:invasion of legume plants by rhizobia. Curr OpinMicrobiol. 3: 613-617.
Gay G, Normand L, Marmeisse R, Sotta B et Debaud JC(1994) Auxin overproducer mutants of Hebelomacylindrosporum Tomagnési have increasedmycorrhizal activity. New Phytol. 128: 645-657.
Gälweiler L, Guan C, Müller A, Wisman E, Mendgen K,Yephremov A et Palme K (1998) Regulation ofpolar auxin transport by AtPIN1 in Arabidopsisvascular tissue. Science. 282: 2226-2230.
Geisler M, Blakeslee JJ, Bouchard R, Lee OR, Vincenzetti V,Bandyopadhyay A, Titapiwatanakun B, Peer WA,Bailly A, Richards EL, Ejendal KF, Smith AP, BarouxC, Grossniklaus U, Müller A, Hrycyna CA, DudlerR, Murphy AS et Martinoia E (2005) Cellular effluxof auxin catalyzed by the Arabidopsis MDR/PGPtransporter AtPGP1. Plant J. 44: 179-194.
Geldner N, Anders N, Wolters H, Keicher J, Kornberger W,Muller P, Delbarre A, Ueda T, Nakano A et JürgensG (2003) The Arabidopsis GNOM ARF-GEF mediatesendosomal recycling, auxin transport, and auxin-dependent plant growth. Cell. 112: 219-230.
Geldner N, Richter S, Vieten A, Marquardt S, Torres-RuizRA, Mayer U et Jürgens G (2004) Partial loss-of-function alleles reveal a role for GNOM in auxintransport-related, post-embryonic development ofArabidopsis. Development. 131: 389-400.
Geurts R, Fedorova E et Bisseling T (2005) Nod factorsignaling genes and their function in the early stagesof Rhizobium infection. Curr Opin Plant Biol. 8: 346-352.
Gonzalez-Rizzo S, Crespi M et Frugier F (2006) TheMedicago truncatula CRE1 Cytokinin ReceptorRegulates Lateral Root Development and EarlySymbiotic Interaction with Sinorhizobium meliloti.Plant Cell. 18: 2680-2693.
Gordons A, Stevens JR et Berry AM (1988) Cytokininsecretion by Frankia sp. HFPArI3 in defined medium.Plant Physiol. 87: 15-16.
Graham PH et Vance CP (2003) Legumes: importance andconstraints to greater use. Plant Physiol. 131: 872-877.
Halbleib CM et Ludden PW (2000) Regulation of biologicalnitrogen fixation. J Nutr. 130: 1081-1084.
Hammad Y, Nalin R, Marechal J, Fiasson K, Pepin R, BerryAM, Normand P et Domenach AM (2003) Apossible role for phenyl acetic acid (PAA) on Alnusglutinosa nodulation by Frankia. Plant Soil. 254:193-205.
Hanahan D (1983) Studies on transformation of Escherichiacoli with plasmids. J Mol Biol. 166: 557-580.
Handberg K et Stougaard J (1992) Lotus japonicus, anautogamous, diploid legume species for classicaland molecular genetics. Plant J. 2: 487-496.
Hardy RWF, et Knight E (1968) The biochemistry andpostulated mechanism of N2 fixation. In Progress inphytochemistry. pp 387-469. L. Reinhold (ed).
Heidstra R, Yang WC, Yalcin Y, Peck S, Emons AM, vanKammen A et Bisseling T (1997) Ethylene providespositional information on cortical cell division but isnot involved in Nod factor-induced root hair tipgrowth in Rhizobium-legume interaction.Development. 124: 1781-1787.
Heinekamp T, Kuhlmann M, Lenk A, Strathmann A etDröge-Laser W (2002) The tobacco bZIPtranscription factor BZI-1 binds to G-box elementsin the promoters of phenylpropanoid pathway genesin vitro, but it is not involved in their regulation invivo. Mol Genet Genomics. 267: 16-26.
Heinekamp T, Strathmann A, Kuhlmann M, Froissard M,Müller A, Perrot-Rechenmann C et Dröge-LaserW (2004) The tobacco bZIP transcription factor BZI-1binds the GH3 promoter in vivo and modulatesauxin-induced transcription. Plant J. 38: 298-309.
Helariutta Y, Fukaki H, Wysocka-Diller J, Nakajima K,Jung J, Sena G, Hauser MT et Benfey PN (2000)The SHORT-ROOT gene controls radial patterning ofthe Arabidopsis root through radial signaling. Cell.101: 555-567.
Henrissat B et Davies GJ (1997) Structural and sequencebased classification of glycosyl hydrolases. Curr OpinStruct Biol. 7: 637-644.
Himanen K, Boucheron E, Vanneste S, de Almeida EnglerJ, Inzé D et Beeckman T (2002) Auxin-mediatedcell cycle activation during early lateral root initiation.Plant Cell. 14: 2339-2351.
Hirsch AM, Lum MR et Downie JA (2001) What makes therhizobia-legume symbiosis so special? Plant Physiol.127: 1484-1492.
Hoagland DR et Arnon DI (1938) Synthetic media forhydroponic culture. Calif Agr Expt Public. 347: 35-37.
Hobbie LJ (2006) Auxin and cell polarity: the emergence ofAXR4. Trends Plant Sci. 11: 517-518.
Hocher V, Auguy F, Argout X, Laplaze L, Franche C etBogusz D (2006) Expressed sequence-tag analysisin Casuarina glauca actinorhizal nodule and root.New Phytol. 169: 681-688.
Howard JB, et Rees DC (2000) Structure of the nitrogenaseprotein components. In Prokaryotic nitrogen fixation: a model system for the analysis of a biologicalprocess. Triplett, EW (ed) Horizon Scientific Press,Wymondham, UK.

66
Hughes M, Donnelly C, Crozier A et Wheeler CT (1999)Effects of the exposure of roots of Alnus glutinosa tolight on flavonoids and nodulation. Can J Bot. 77:1311-1315.
Huss-Danel K (1997) Actinorhizal symbiosis and their N2fixation. New Phytol. 136: 375-405.
Hutcheson SW et Kosuge T (1985) Regulation of 3-indoleacetic acid production in Pseudomonassyringae pv. savastanoi. Purification and propertiesof tryptophan 2-monooxygenase. J Biol Chem. 260:6281-6287.
Jasinski M, Ducos E, Martinoia E et Boutry M (2003) TheATP-binding cassette transporters: structure,function, and gene family comparison between riceand Arabidopsis. Plant Physiol. 131: 1169-1177.
Jentschel K, Thiel D, Rehn F et Ludwig-Müller J (2006)Arbuscular mycorrhiza enhances auxin levels andalters auxin biosynthesis in Tropaeolum majusduring early stage of colonization. Physiol Plant. 129:320-333.
Kaló P, Gleason C, Edwards A, Marsh J, Mitra RM, Hirsch S,Jakab J, Sims S, Long SR, Rogers J, Kiss GB,Downie JA et Oldroyd GE (2005) Nodulationsignaling in legumes requires NSP2, a member ofthe GRAS family of transcriptional regulators.Science. 308: 1786-1789.
Kaneshiro T et Kwolek WF (1985) Stimulated nodulation ofsoybeans by Rhizobium japonicum mutant (B-14075) that catolizes the conversion of tryptophanto indol-3yl-acetic acid. Plant Science. 42: 141-146.
Kape R, Parniske M et Werner D (1991) Chemotaxis andnod Gene Activity of Bradyrhizobium japonicum inResponse to Hydroxycinnamic Acids andIsoflavonoids. Appl Environ Microbiol. 57: 316-319.
Kepinski S et Leyser O (2005a) Plant development: auxin inloops. Curr Biol. 15: R208-R210.
Kepinski S et Leyser O (2005b) The Arabidopsis F-boxprotein TIR1 is an auxin receptor. Nature. 435: 446-451.
Kerr ID et Bennett MJ (2007) New insight into thebiochemical mechanisms regulating auxin transportin plants. Biochem J. 401: 613-622.
Kim HB, Lee H, Oh CJ, Lee NH et An CS (2007) Expressionof EuNOD-ARP1 Encoding Auxin-repressed ProteinHomolog Is Upregulated by Auxin and Localized tothe Fixation Zone in Root Nodules of Elaeagnusumbellata. Mol Cells. 23: 115-121.
Kondorosi E, et Kondorosi A (2000) Control of root noduleorganogenesis. In Prokaryotic nitrogen fixation : amodel system for the analysis of a biologicalprocess. Triplett, EW (ed) Horizon Scientific Press,Wymondham, UK.
Kowalczyk M et Sandberg G (2001) Quantitative analysis ofindole-3-acetic acid metabolites in Arabidopsis.Plant Physiol. 127: 1845-1853.
Kramer EM et Bennett MJ (2006) Auxin transport: a field influx. Trends Plant Sci. 11: 382-386.
Ky CL, Barre P, Lorieux M, Trouslot P, Akaffou S, Louarn J,Charrier A, Hamon S et Noirot M (2000)Interspecific genetic linkage map, segregationdistortion and genetic conversion in coffee (Coffeasp.). Theor Appl Genet. 101: 669-676.
Lambers H, Shane MW, Cramer MD, Pearse SJ etVeneklaas EJ (2006) Root structure and functioning
for efficient acquisition of phosphorus: Matchingmorphological and physiological traits. Ann Bot(Lond). 98: 693-713.
Lambrecht M, Okon Y, Vande Broek A et Vanderleyden J(2000) Indole-3-acetic acid: a reciprocal signallingmolecule in bacteria-plant interactions. TrendsMicrobiol. 8: 298-300.
Laplaze L (1999) Approche moléculaire de la mise en placede la symbiose actinorhizienne. Thèse.
Laplaze L, Duhoux E, Franche C, Frutz T, Svistoonoff S,Bisseling T, Bogusz D et Pawlowski K (2000a)Casuarina glauca prenodule cells display the samedifferentiation as the corresponding nodule cells.Mol Plant Microbe Interact. 13: 107-112.
Laplaze L, Parizot B, Baker A, Ricaud L, Martinière A,Auguy F, Franche C, Nussaume L, Bogusz D etHaseloff J (2005) GAL4-GFP enhancer trap lines forgenetic manipulation of lateral root development inArabidopsis thaliana. J Exp Bot. 56: 2433-2442.
Laplaze L, Ribeiro A, Franche C, Duhoux E, Auguy F,Bogusz D et Pawlowski K (2000b) Characterizationof a Casuarina glauca nodule-specific subtilisin-likeprotease gene, a homolog of Alnus glutinosa ag12.Mol Plant Microbe Interact. 13: 113-117.
Laskowski M, Biller S, Stanley K, Kajstura T et Prusty R(2006) Expression Profiling of Auxin-TreatedArabidopsis Roots: Toward a Molecular Analysis ofLateral Root Emergence. Plant Cell Physiol. 47: 788-792.
Lee KH et Larue TA (1992) Exogenous Ethylene InhibitsNodulation of Pisum sativum L. cv Sparkle. PlantPhysiol. 100: 1759-1763.
Leyser O (2006) Dynamic integration of auxin transport andsignalling. Curr Biol. 16: R424-R433.
Limpens E, Franken C, Smit P, Willemse J, Bisseling T etGeurts R (2003) LysM domain receptor kinasesregulating rhizobial Nod factor-induced infection.Science. 302: 630-633.
Ljung K, Bhalerao RP et Sandberg G (2001) Sites andhomeostatic control of auxin biosynthesis inArabidopsis during vegetative growth. Plant J. 28:465-474.
Ljung K, Hull AK, Celenza J, Yamada M, Estelle M,Normanly J et Sandberg G (2005) Sites andregulation of auxin biosynthesis in Arabidopsisroots. Plant Cell. 17: 1090-1104.
Ljung K, Hull AK, Kowalczyk M, Marchant A, Celenza J,Cohen JD et Sandberg G (2002) Biosynthesis,conjugation, catabolism and homeostasis of indole-3-acetic acid in Arabidopsis thaliana. Plant Mol Biol.49: 249-272.
Lohar DP, Schaff JE, Laskey JG, Kieber JJ, Bilyeu KD et BirdDM (2004) Cytokinins play opposite roles in lateralroot formation, and nematode and Rhizobialsymbioses. Plant J. 38: 203-214.
Madsen EB, Madsen LH, Radutoiu S, Olbryt M, RakwalskaM, Szczyglowski K, Sato S, Kaneko T, Tabata S,Sandal N et Stougaard J (2003) A receptor kinasegene of the LysM type is involved in legumeperception of rhizobial signals. Nature. 425: 637-640.
Malamy JE et Benfey PN (1997) Organization and celldifferentiation in lateral roots of Arabidopsisthaliana. Development. 124: 33-44.

67
Manulis S, Haviv-Chesner A, Brandl MT, Lindow SE etBarash I (1998) Differential involvement of indole-3-acetic acid biosynthetic pathways in pathogenicityand epiphytic fitness of Erwinia herbicola pv.gypsophilae. Mol Plant Microbe Interact. 11: 634-642.
Marchant A, Bhalerao R, Casimiro I, Eklöf J, Casero PJ,Bennett M et Sandberg G (2002) AUX1 promoteslateral root formation by facilitating indole-3-aceticacid distribution between sink and source tissues inthe Arabidopsis seedling. Plant Cell. 14: 589-597.
Marchant A, Kargul J, May ST, Muller P, Delbarre A,Perrot-Rechenmann C et Bennett MJ (1999) AUX1regulates root gravitropism in Arabidopsis byfacilitating auxin uptake within root apical tissues.EMBO J. 18: 2066-2073.
Marín-Rodríguez MC, Orchard J et Seymour GB (2002)Pectate lyases, cell wall degradation and fruitsoftening. J Exp Bot. 53: 2115-2119.
Martinoia E, Klein M, Geisler M, Bovet L, Forestier C,Kolukisaoglu U, Müller-Röber B et Schulz B(2002) Multifunctionality of plant ABC transporters--more than just detoxifiers. Planta. 214: 345-355.
Mateos PF, Baker DL, Petersen M, Velázquez E, Jiménez-Zurdo JI, Martínez-Molina E, Squartini A,Orgambide G, Hubbell DH et Dazzo FB (2001)Erosion of root epidermal cell walls by Rhizobiumpolysaccharide-degrading enzymes as related toprimary host infection in the Rhizobium-legumesymbiosis. Can J Microbiol. 47: 475-487.
Mathesius U (2001) Flavonoids induced in cells undergoingnodule organogenesis in white clover are regulatorsof auxin breakdown by peroxidase. J Exp Bot. 52:419-426.
Mathesius U, Charon C, Rolfe BG, Kondorosi A et CrespiM (2000a) Temporal and spatial order of eventsduring the induction of cortical cell divisions in whiteclover by Rhizobium leguminosarum bv. trifoliiinoculation or localized cytokinin addition. Mol PlantMicrobe Interact. 13: 617-628.
Mathesius U, Schlaman HR, Spaink HP, Of Sautter C, RolfeBG et Djordjevic MA (1998) Auxin transportinhibition precedes root nodule formation in whiteclover roots and is regulated by flavonoids andderivatives of chitin oligosaccharides. Plant J. 14: 23-34.
Mathesius U, Weinman JJ, Rolfe BG et Djordjevic MA(2000b) Rhizobia can induce nodules in white cloverby "hijacking" mature cortical cells activated duringlateral root development. Mol Plant Microbe Interact.13: 170-182.
Miller IM et Baker DD (1985) The initiation, developmentand structure of root nodules in Elaegnusangustifolia L. (Elaeagnaceae). Protoplasma. 128:107-119.
Mitra RM, Gleason CA, Edwards A, Hadfield J, Downie JA,Oldroyd GE et Long SR (2004) ACa2+/calmodulin-dependent protein kinaserequired for symbiotic nodule development: Geneidentification by transcript-based cloning. Proc NatlAcad Sci U S A. 101: 4701-4705.
Miwa H, Sun J, Oldroyd GE et Downie JA (2006) Analysis ofNod-factor-induced calcium signaling in root hairsof symbiotically defective mutants of Lotusjaponicus. Mol Plant Microbe Interact. 19: 914-923.
Moiroud A (1996) Diversité et écologie des plantesactinorhiziennes. Acta Bot Gall. 143: 651-661.
Moulin L, Munive A, Dreyfus B et Boivin-Masson C (2001)Nodulation of legumes by members of the beta-subclass of Proteobacteria. Nature. 411: 948-950.
Möller S, Croning MD et Apweiler R (2001) Evaluation ofmethods for the prediction of membrane spanningregions. Bioinformatics. 17: 646-653.
Mulder L, Hogg BV, Bersoult A et Cullimore JV (2005)Integration of signalling pathways in theestablishment of the legume-rhizobia symbiosis.Physiologia Plantarum. 123: 207-218.
Mullin BC et Dobritsa SV (1996) Molecular analysis ofactinorhizal symbiotic systems : progress to date.Plant Soil. 186: 9-20.
Murashige T et Skoog F (1962) A revised medium for rapidgrowth and bioassays with tobacco tissue cultures.Physiol Plant. 15: 473-497.
Navarro L, Dunoyer P, Jay F, Arnold B, Dharmasiri N,Estelle M, Voinnet O et Jones JD (2006) A plantmiRNA contributes to antibacterial resistance byrepressing auxin signaling. Science. 312: 436-439.
Neumann G et Martinoia E (2002) Cluster roots--anunderground adaptation for survival in extremeenvironments. Trends Plant Sci. 7: 162-167.
Neuteboom LW, Ng JM, Kuyper M, Clijdesdale OR,Hooykaas PJ et van der Zaal BJ (1999a) Isolationand characterization of cDNA clones correspondingwith mRNAs that accumulate during auxin-inducedlateral root formation. Plant Mol Biol. 39: 273-287.
Neuteboom LW, Veth-Tello LM, Clijdesdale OR, HooykaasPJ et van der Zaal BJ (1999b) A novel subtilisin-likeprotease gene from Arabidopsis thaliana isexpressed at sites of lateral root emergence. DNARes. 6: 13-19.
Noh B, Murphy AS et Spalding EP (2001) Multidrugresistance-like genes of Arabidopsis required forauxin transport and auxin-mediated development.Plant Cell. 13: 2441-2454.
Normand P, Lapierre P, Tisa LS, Gogarten JP, Alloisio N,Bagnarol E, Bassi CA, Berry AM, Bickhart DM,Choisne N, Couloux A, Cournoyer B, Cruveiller S,Daubin V, Demange N, Francino MP, Goltsman E,Huang Y, Kopp OR, Labarre L, Lapidus A, Lavire C,Marechal J, Martinez M, Mastronunzio JE, MullinBC, Niemann J, Pujic P, Rawnsley T, Rouy Z,Schenowitz C, Sellstedt A, Tavares F, Tomkins JP,Vallenet D, Valverde C, Wall LG, Wang Y, MedigueC et Benson DR (2006) Genome characteristics offacultatively symbiotic Frankia sp. strains reflect hostrange and host plant biogeography. Genome Res.17: 7-15.
Nukui N, Ezura H, Yuhashi K, Yasuta T et Minamisawa K(2000) Effects of ethylene precursor and inhibitorsfor ethylene biosynthesis and perception onnodulation in Lotus japonicus and Macroptiliumatropurpureum. Plant Cell Physiol. 41: 893-897.
Obertello M, Santi C, Sy MO, Laplaze L, Auguy F, Bogusz Det Franche C (2005) Comparison of four constitutivepromoters for the expression of transgenes in thetropical nitrogen-fixing tree Allocasuarina verticillata.Plant Cell Rep. 24: 540-548.
Obertello M, Sy MO, Laplaze L, Santi C, Svistoonoff S,Auguy F, Bogusz D et Franche C (2003)Actinorhizal nitrogen fixing nodules : infection

68
process, molecular biology and genomics. Afr JBiotechnol. 2: 528-538.
Okushima Y, Fukaki H, Onoda M, Theologis A et TasakaM (2007) ARF7 and ARF19 Regulate Lateral RootFormation via Direct Activation of LBD/ASL Genes inArabidopsis. Plant Cell. 19: 118-130.
Oldroyd GE et Downie JA (2006) Nuclear calcium changesat the core of symbiosis signalling. Curr Opin PlantBiol. 9: 351-357.
Ovtsyna AO, Dolgikh EA, Kilanova AS, Tsyganov VE,Borisov AY, Tikhonovich IA et Staehelin C (2005)Nod factors induce nod factor cleaving enzymes inpea roots. Genetic and pharmacological approachesindicate different activation mechanisms. PlantPhysiol. 139: 1051-1064.
Paciorek T et Friml J (2006) Auxin signaling. J Cell Sci. 119:1199-1202.
Pacios-Bras C, Schlaman HR, Boot K, Admiraal P,Langerak JM, Stougaard J et Spaink HP (2003)Auxin distribution in Lotus japonicus during rootnodule development. Plant Mol Biol. 52: 1169-1180.
Paponov IA, Teale WD, Trebar M, Blilou I et Palme K(2005) The PIN auxin efflux facilitators: evolutionaryand functional perspectives. Trends Plant Sci. 10:170-177.
Parry G, Delbarre A, Marchant A, Swarup R, Napier R,Perrot-Rechenmann C et Bennett MJ (2001a)Novel auxin transport inhibitors phenocopy theauxin influx carrier mutation aux1. Plant J. 25: 399-406.
Parry G, Marchant A, May S, Swarup R, Swarup K, James N,Graham N, Allen T, Martucci T, Yemm A, Napier R,Manning K, King G et Bennett M (2001b) Quick onthe uptake : Characterization of a family of plantauxin influx carriers. J Plant Growth Reg. 20: 217-225.
Patriarca EJ, Tatè R, Ferraioli S et Iaccarino M (2004)Organogenesis of legume root nodules. Int RevCytol. 234: 201-262.
Pawlowski K et Bisseling T (1996) Rhizobial andActinorhizal Symbioses: What Are the SharedFeatures? Plant Cell. 8: 1899-1913.
Pawlowski K et Sirrenberg A (2003) Symbiosis betweenFrankia and actinorhizal plants: root nodules of non-legumes. Indian J Exp Biol. 41: 1165-1183.
Perret X, Staehelin C et Broughton WJ (2000) Molecularbasis of symbiotic promiscuity. Microbiol Mol BiolRev. 64: 180-201.
Petrasek J, Mravec J, Bouchard R, Blakeslee JJ, Abas M,Seifertová D, Wisniewska J, Tadele Z, Kubes M,Covanová M, Dhonukshe P, Skupa P, Benková E,Perry L, Krecek P, Lee OR, Fink GR, Geisler M,Murphy AS, Luschnig C, Zazimalova E et Friml J(2006) PIN Proteins Perform a Rate-Limiting Functionin Cellular Auxin Efflux. Science. 312: 914-918.
Prayitno J, Rolfe BG et Mathesius U (2006) The EthyleneInsensitive sickle Mutant of Medicago truncatulaShows Altered Auxin Transport Regulation duringNodulation. Plant Physiol. 142: 168-180.
Quint M et Gray WM (2006) Auxin signaling. Curr OpinPlant Biol. 8: 448-453.
Radutoiu S, Madsen LH, Madsen EB, Felle HH, Umehara Y,Grønlund M, Sato S, Nakamura Y, Tabata S, SandalN et Stougaard J (2003) Plant recognition of
symbiotic bacteria requires two LysM receptor-likekinases. Nature. 425: 585-592.
Rangaraj P, Rüttimann-Johnson C, Shah VK, et LuddenPW (2000) Biosynthesis of the iron-molybdenumand iron-vanadium cofactors of the nif- and vnf-encoded nitrogenases. In Prokaryotic nitrogenfixation : a model system for the analysis of abiological process. Triplett, EW (ed) Horizon ScientificPress, Wymondham, UK.
Rashotte AM, Brady SR, Reed RC, Ante SJ et Muday GK(2000) Basipetal auxin transport is required forgravitropism in roots of Arabidopsis. Plant Physiol.122: 481-490.
Reed RC, Brady SR et Muday GK (1998) Inhibition of auxinmovement from the shoot into the root inhibitslateral root development in Arabidopsis. PlantPhysiol. 118: 1369-1378.
Rees DC et Howard JB (2000) Nitrogenase: standing at thecrossroads. Curr Opin Chem Biol. 4: 559-566.
Rubio LM et Ludden PW (2005) Maturation of nitrogenase:a biochemical puzzle. J Bacteriol. 187: 405-414.
Sauer M, Balla J, Luschnig C, Wisniewska J, Reinöhl V,Friml J et Benková E (2006) Canalization of auxinflow by Aux/IAA-ARF-dependent feedbackregulation of PIN polarity. Genes Dev. 20: 2902-2911.
Savka MA, Dessaux Y, Oger P et Rossbach S (2002)Engineering bacterial competitiveness andpersistence in the phytosphere. Mol Plant MicrobeInteract. 15: 866-874.
Schmidt JS, Harper JE, Hoffman TK et Bent AF (1999)Regulation of soybean nodulation independent ofethylene signaling . Plant Physiol. 119: 951-960.
Schnabel EL et Frugoli J (2004) The PIN and LAX families ofauxin transport genes in Medicago truncatula. MolGenet Genomics. 272: 420-432.
Schwencke J, Bureau JM, Crosnier MT et Brown S (1998)Cytometric determination of genome size and basecomposition of tree species of three genera ofCasuarinaceae. Plant Cell Rep. 18: 346-349.
Smit P, Raedts J, Portyanko V, Debellé F, Gough C,Bisseling T et Geurts R (2005) NSP1 of the GRASprotein family is essential for rhizobial Nod factor-induced transcription. Science. 308: 1789-1791.
Soltis DE, Soltis PS, Morgan DR, Swensen SM, Mullin BC,Dowd JM et Martin PG (1995) Chloroplast genesequence data suggest a single origin of thepredisposition for symbiotic nitrogen fixation inangiosperms. Proc Natl Acad Sci U S A. 92: 2647-2651.
Stacey G, Libault M, Brechenmacher L, Wan J et May GD(2006) Genetics and functional genomics of legumenodulation. Curr Opin Plant Biol. 9: 110-121.
Supanjani S, Habib A, Mabood F, Lee KD, Donnelly D etSmith DL (2006) Nod factor enhances calciumuptake by soybean. Plant Physiol Biochem. 44: 866-872.
Svistoonoff S (2003) Implication d'une subtilase dans lesétapes précoces des symbioses actinorhiziennes.Thèse.
Svistoonoff S, Laplaze L, Auguy F, Runions J, DuponnoisR, Haseloff J, Franche C et Bogusz D (2003) cg12expression is specifically linked to infection of roothairs and cortical cells during Casuarina glauca and

69
Allocasuarina verticillata actinorhizal noduledevelopment. Mol Plant Microbe Interact. 16: 600-607.
Swarup R et Bennett MJ (2003) Auxin transport: thefountain of life in plants? Dev Cell. 5: 824-826.
Swarup R, Friml J, Marchant A, Ljung K, Sandberg G,Palme K et Bennett MJ (2001) Localization of theauxin permease AUX1 suggests two functionallydistinct hormone transport pathways operate in theArabidopsis root apex. Genes Dev. 15: 2648-2653.
Swarup R, Kargul J, Marchant A, Zadik D, Rahman A, MillsR, Yemm A, May S, Williams L, Millner P, TsurumiS, Moore I, Napier R, Kerr ID et Bennett MJ (2004)Structure-function analysis of the presumptiveArabidopsis auxin permease AUX1. Plant Cell. 16:3069-3083.
Swarup R, Kramer EM, Perry P, Knox K, Leyser HM,Haseloff J, Beemster GT, Bhalerao R et Bennett MJ(2005) Root gravitropism requires lateral root capand epidermal cells for transport and response to amobile auxin signal. Nat Cell Biol. 7: 1057-1065.
Swensen SM (1996) The evolution of actinorhizal symbiosis:evidence for multiple origins of the symbioticassociation. Am J Bot. 83: 1503-1512.
Teale WD, Paponov IA et Palme K (2006) Auxin in action:signalling, transport and the control of plant growthand development. Nat Rev Mol Cell Biol. 7: 847-859.
Terasaka K, Blakeslee JJ, Titapiwatanakun B, Peer WA,Bandyopadhyay A, Makam SN, Lee OR, RichardsEL, Murphy AS, Sato F et Yazaki K (2005) PGP4, anATP binding cassette P-glycoprotein, catalyzes auxintransport in Arabidopsis thaliana roots. Plant Cell.17: 2922-2939.
Tian Q, Uhlir NJ et Reed JW (2002) Arabidopsis SHY2/IAA3inhibits auxin-regulated gene expression. Plant Cell.14: 301-319.
Timmers AC, Auriac MC et Truchet G (1999) Refinedanalysis of early symbiotic steps of the Rhizobium-Medicago interaction in relationship withmicrotubular cytoskeleton rearrangements.Development. 126: 3617-3628.
Timmers AC, Vallotton P, Heym C et Menzel D (2007)Microtubule dynamics in root hairs of Medicagotruncatula. Eur J Cell Biol. 86: 69-83.
Tirichine L, Sandal N, Madsen LH, Radutoiu S, AlbrektsenAS, Sato S, Asamizu E, Tabata S et Stougaard J(2007) A gain-of-function mutation in a cytokininreceptor triggers spontaneous root noduleorganogenesis. Science. 315: 104-107.
Torrey JG (1976) Initiation and development of root nodulesof Casuarina (Casuarinaceae). Am J Bot. 63: 335-345.
Tyagi AK, Khurana JP, Khurana P, Raghuvanshi S, Gaur A,Kapur A, Gupta V, Kumar D, Ravi V, Vij S, KhuranaP et Sharma S (2004) Structural and functionalanalysis of rice genome. J Genet. 83: 79-99.
Udvardi MK, Tabata S, Parniske M et Stougaard J (2005)Lotus japonicus: legume research in the fast lane.Trends Plant Sci. 10: 222-228.
van Brussel AA, Bakhuizen R, Van Sprosen PC, Spaink HP,Tak T et Lugtenberg BJJ (1992) Induction ofpreinfection thread structures in the leguminoushost plant by mitogenic lipo-oligosaccharides ofRhizobium. Science. 257: 70-71.
van Brussel AA, Recourt K, Pees E, Spaink HP, Tak T,Wijffelman CA, Kijne JW et Lugtenberg BJ (1990) Abiovar-specific signal of Rhizobium leguminosarumbv. viciae induces increased nodulation gene-inducing activity in root exudate of Vicia sativasubsp. nigra. J Bacteriol. 172: 5394-5401.
van Ghelue M, Lovaas E, Ringo E et Solheim B (1997)Production and specificity of root hair deformationfactor(s). Physiol Plant. 99: 579-587.
Vanneste S, De Rybel B, Beemster GT, Ljung K, De Smet I,Van Isterdael G, Naudts M, Iida R, Gruissem W,Tasaka M, Inzé D, Fukaki H et Beeckman T(2005a) Cell cycle progression in the pericycle is notsufficient for SOLITARY ROOT/IAA14-mediatedlateral root initiation in Arabidopsis thaliana. PlantCell. 17: 3035-3050.
Vanneste S, Maes L, De Smet I, Himanen K, Naudts M,Inzé D et Beeckman T (2005b) Auxin regulation ofcell cycle and its role during lateral root initiation.Physiol Plant. 123: 139-146.
Vieten A, Vanneste S, Wisniewska J, Benková E, BenjaminsR, Beeckman T, Luschnig C et Friml J (2005)Functional redundancy of PIN proteins isaccompanied by auxin-dependent cross-regulationof PIN expression. Development. 132: 4521-4531.
Vissenberg K, Fry SC, Pauly M, Höfte H et Verbelen JP(2005) XTH acts at the microfibril-matrix interfaceduring cell elongation. J Exp Bot. 56: 673-683.
Wais RJ, Keating DH et Long SR (2002) Structure-functionanalysis of nod factor-induced root hair calciumspiking in Rhizobium-legume symbiosis. PlantPhysiol. 129: 211-224.
Wasson AP, Pellerone FI et Mathesius U (2006) Silencingthe Flavonoid Pathway in Medicago truncatulaInhibits Root Nodule Formation and Prevents AuxinTransport Regulation by Rhizobia. Plant Cell. 18:1617-1629.
Wen F, Laskowski M et Hawes M (2006) Cell separation inroots. Ann Plant Reviews. 25: 91-105.
Wheeler CT, Crozier A et Sandberg G (1984) Thebiosynthesis of indole-3-acetic acid by Frankia. PlantSoil. 78: 99-104.
Wheeler CT, Henson IE et MacLaughlin ME (1979)Hormones in plants bearing actinomycete nodules.Bot Gaz. 140: 52-57.
Wisniewska J, Xu J, Seifertová D, Brewer PB, Ruzicka K,Blilou I, Roquie D, Benková E, Scheres B et Friml J(2006) Polar PIN Localization Directs Auxin Flow inPlants. Science. 312: 883.
Wood SM et Newcomb W (1989) Nodule morphogenesis:the early infection of Alfalfa (Medicago sativa) roothairs by Rhizobium meliloti. Can J Bot. 67: 3108-3122.
Woodward AW et Bartel B (2005) Auxin: regulation, action,and interaction. Ann Bot (Lond). 95: 707-735.
Yang WC, Canter Cremers HC, Hogendijk P, Katinakis P,Wijffelman CA, Franssen H, van Kammen A etBisseling T (1992) In-situ localization of chalconesynthase mRNA in pea root nodule development.Plant J. 2: 143-151.
Yang Y, Hammes UZ, Taylor CG, Schachtman DP etNielsen E (2006) High-Affinity Auxin Transport bythe AUX1 Influx Carrier Protein. Curr Biol. 160:1123-1127.

70
Young GB, Jack DL, Smith DW et Saier MH (1999) Theamino acid/auxin:proton symport permease family.Biochim Biophys Acta. 1415: 306-322.
Zhang B, Wang Q et Pan X (2007) MicroRNAs and theirregulatory roles in animals and plants. J Cell Physiol.210: 279-289.

RÉSUMÉLes plantes actinorhiziennes appartiennent à 8 familles d’angiosperme et forment une symbiose fixatrice
d’azote avec l’actinomycète du sol Frankia qui aboutit à la formation de nodules au niveau du système racinaire dela plante. Le nodule actinorhizien est considéré comme une racine latérale modifiée car i) il provient de divisions descellules du péricycle situées en face du pôle de xylème, ii) il possède un méristème apical et un système vasculairecentral et iii) chez certaines espèces comme Casuarina glauca une racine nodulaire est produite à l’apex de chaquelobe nodulaire. L’auxine, et notamment le transport d’influx, est impliquée dans la mise en place de la racinelatérale. Nous avons donc identifié des gènes de transporteurs d’influx d’auxine chez la plante actinorhizienne C.glauca et étudié le rôle du transport d’influx au cours de la mise en place du nodule actinorhizien.
Deux gènes de la famille AUX-LAX codant des transporteurs d’influx d’auxine ont été identifiés C. glauca. Lesprofils d’expression des gènes CgAUX1 et CgLAX3 sont très conservés entre C. glauca et Arabidopsis thaliana. Deplus, des analyses fonctionnelles par complémentation de mutants d’A. thaliana ont mis en évidence uneéquivalence entre CgAUX1 et AtAUX1. Nos études suggèrent également qu’il existe une divergence fonctionnelle ausein de la famille AUX-LAX.
Nous avons analysé le rôle de ces gènes au cours de la mise en place de la symbiose. Notre étude montre que legène CgAUX1 est exprimé dans les cellules infectées tout au long de l’infection. De plus, le rôle du transport d’influxd’auxine dans le mécanisme d’infection a été confirmé par l’utilisation d’un inhibiteur du transport d’influx. Parailleurs, le gène CgAUX1 est exprimé dans le primordium de racine latérale mais pas dans le primordium nodulaire.Cela suggère que ces deux organes présentent des différences dans leur programme de développement.
Afin d’identifier les mécanismes agissant en aval du transport d’influx d’auxine, nous avons étudié le rôled’AtLAX3 chez Arabidopsis. Nous avons montré qu’un certain nombre de gènes de remodelage de la paroi sontinduits par l’auxine de façon dépendante d’AtLAX3 au cours de l’émergence de la racine latérale. Nous avonscherché à identifier des gènes de remodelage de la paroi qui pourraient être impliqués dans l’infection par labactérie Frankia de façon dépendante de CgAUX1. Cg12 qui code une protéase de type subtilisine spécifiquementexprimée dans les cellules infectées pourrait être une cible de la signalisation auxinique dépendante de CgAUX1.
Nos résultats suggèrent que le transport d’influx d’auxine est impliqué dans la mise en place du noduleactinorhizien chez C. glauca.
ABSTRACTActinorhizal plants belonging to 8 families of angiosperms can enter symbiosis with a soil actinomycete called
Frankia. This interaction leads to the formation of nitrogen fixing nodules on the plant root system. The actinorhizalnodule is considered as a modified lateral root because i) it originates from divisions of pericycle cells situated infront of xylem poles, ii) its vasculature is central and its growth is indeterminate due to the presence of an apicalmeristem and iii) in some species such as Casuarina glauca a so-called “nodular root” is formed at the apex of eachnodule lobe. Auxin, and more particularly auxin influx, is involved in lateral root formation. We identified auxininflux transporter genes in the actinorhizal plant C. glauca and studied the role of auxin influx transport duringactinorhizal nodule formation.
Two AUX-LAX genes encoding for auxin influx carriers have been identified in C. glauca. The expressionpatterns of CgAUX1 and CgLAX3 are highly conserved between C. glauca and Arabidopsis thaliana. Functionalcomplementation of the Arabidopsis aux1 mutant revealed that CgAUX1 and AtAUX1 share equivalent functions.Our data suggest that functional divergence exists in the AUX-LAX family.
We analysed the role of these genes during the actinorhizal symbiosis. Expression studies showed that CgAUX1is expressed in all infected cells. Moreover, we confirmed that auxin influx transport is involved in the symbioticprocess by taking advantage of an auxin influx transport inhibitor. We also observed that CgAUX1 is expressed inlateral root primordium but not in nodule primordium thus pinpointing some differences in the developmentalprogram of these two organs.
We then tried to identify the mechanisms acting downstream of auxin influx transport by studying the role ofAtLAX3 in Arabidopsis. We showed that a set of cell wall remodeling genes are induced by auxin in a AtLAX3dependent way during lateral root emergence. We next tried to identify cell wall remodeling genes that could beinvolved in the infection process in a CgAUX1 dependent way. Cg12 encodes for a subtilisin-like protease that isspecifically expressed in Frankia infected cells and could be a target of CgAUX1 dependent auxin signaling.
Our results suggest that auxin influx transport is involved in the infection process during actinorhizal noduleformation in C. glauca.





























