Embed Size (px)
Citation preview

332
BBA 65326
BIOCHIMICA ET BIOPHYSICA ACTA
ADENOSINE TRIPHOSPHATASE ACTIVITY OF ADRENAL MEDULLACATECHOLAMINE GRANULES
N. KIRSHNER, A. G. KIRSHNER AND D. L. KAMIN
Departments oj Biochemistry and Experimental Surgery, Duke University Medical Center, Dwrham,N.C. (U.S.A.j
(Received May 28th, 1965)
SUMMARY
The catecholamine-containing granules of cow-adrenal medulla were separatedfrom mitochondria by centrifugation through a linear continuous sucrose densitygradient in a swinging-bucket rotor. Fractions collected by puncturing the bottomof the centrifuge tube were assayed for catecholamines, total and insoluble protein,succinate-dehydrogenase (succinate: (acceptor) oxidoreductase, EC 1.3.99.I) activityand adenosine triphosphatase (ATP phosphohydrolase, EC 3.6.1.3) activity. Thepresence of an adenosine-triphosphatase activity which closely paralleled the distribution of catecholamine granules and the inhibition of this adenosine triphosphatase byN-methylmaleimide suggests a role for this enzyme in the uptake of catecholamines.
INTRODUCTION
The catecholamine-containing granules of the adrenal medulla isolated inisotonic sucrose can take up adrenaline from the external medium at 300 but not at 00
(refs. 1,2). This uptake is stimulated by ATP plus Mg2+ and inhibited by EDTA,reserpine and other drugs and by some reagents which react with sulfhydryl groups'.".Pre-incubation of the granules for IO min with O.I mM N-ethylmaleimide completelyinhibited the Mg2+ plus ATP-stimulated uptake. However, this inhibition was prevented ifthe pre-incubation of granules with N-ethylmaleimide was carried out in thepresence of Mg2+ plus ATp3. It was also observed that N-ethylmaleimide inhibitedthe adenosine-triphosphatase (ATP phosphohydrolase, EC 3.6.I.3) activity associatedwith these granule preparations and suggested that this adenosine-triphosphataseactivity might be involved in the stimulated uptake of adrenaline",
The granule preparations used in the above experiments were contaminatedwith mitochondria. However, HILLARp 4 , using a similarly prepared granule preparation, was able to separate the catecholamine-containing granule from the mitochondria by centrifugation through a discontinuous sucrose density gradient and hadshown that the granules contained an adenosine-triphosphatase activity which wasnot due to mitochondrial contamination. In contradiction to these observations,
Biochim, Biophys. Acta, II3 (1966) 332-335

ATPASE ACTIVITY OF CATECHOLAMINE GRANULES 333
FORTIER et at.6 and HAGEN8 were unable to obtain granules entirely free of mitochondria, and concluded that the adenosine-triphosphatase activity in the granulefraction was probably due to mitochondrial contamination. Because of the possiblerelevance of the adenosine-triphosphatase activity to the uptake or secretion ofcatecholamines we have reinvestigated and confirmed the presence of this enzyme inthe catecholamine granules of beef-adrenal medulla.
METHODS
Preparation of gran-ule fractionsThe large granule fractions, relatively free of rnitochondria-, were prepared as
previously described" and further fractionated by centrifugation through a continuouslinear sucrose density gradient. 3 ml of a granule suspension containing 300-500 mgof granules in 0,25 M sucrose were layered over 30 ml of the sucrose gradient varyingin concentration from 1.0 to 2.25 M and centrifuged at 25 000 rev.jrnin for 4 h in aSpinco SW-25 swinging-bucket rotor. Fractions of I.S ml were collected by puncturingthe bottom of the tube, and each was diluted to 20 ml with distilled water to lyse thegranules. A I-ml aliquot was removed for assay of catecholamines" and a o.r-mlaliquot for assay of total proteins. The remainder was diluted to 40 ml with distilledwater and centrifuged at 25 000 X g for 30 min. The clear supernatants were discardedand the residues suspended in 3 ml of distilled water using a Potter-Elvehjem homogenizer. A o.r-ml aliquot was removed for assay of insoluble protein" and o.y-mlaliquots were used for measurement of adenosine-triphosphatase and succinate-dehydrogenase (succinate: (acceptor) oxidoreductase, EC 1.3.99.1) activities.
A denosine-triphosphatase activityThe adenosine-triphosphatase activity of each of the fractions was determined
in the presence and absence of N-ethylmaleimide by measuring the amount of Piliberated upon incubation with ATP. A o.y-ml aliquot of each fraction was pre-incubated for 10 min at 37° with 0.1 ml of 0.5 M Tris buffer (pH 7.4), 0.1 ml of 0.005 MMgS04J and either o. I ml of H20 or 0,I ml of 2 mM N-ethylmaleimide. The reactionwas initiated by the addition of 0.1 ml of 0.05 M ATP (pH 7-4) and allowed to continuefor 30 min at 37°. The incubation was terminated by the addition of La ml of cold 10 %trichloroacetic acid, and after centrifugation, the supernatants were assayed for Pi(ref. 9).
5$tccinate-dehydrogenase activityThe succinate-dehydrogenase activity was determined by the method Of'vVILLI
AMS et at. l O, A 0.5 ml-aliquot of each fraction was incubated for 30 min at 37° withI ml of 0.2 M phosphate buffer (pH 7.4), 0.1 ml of 0.1 mM cytochrome c, 0,1 mlofneotetrazolium chloride (5 mg/mll. and 0.2 ml of 0.2 M sodium succinate (pH 7.4).The reaction was terminated by the addition of 3 ml of acetone, and after centrifugation, the absorbance at 500 mf.l was measured.
RESULTS AND DISCUSSION
The analyses of the fractions obtained from one of the centrifuge tubes are
Biochim. Biopliys, Acta, II3 (1966) 332-335
334
8
7
"C
~gO.2Q;0:
0::-
~ 0.1E
""
II Catechol Amines• Tatal Proteinso Insoluble Proteins
N. KIRSHNER, A. G. KIRSHNER, D. L. KAMIN
04 ~
O.3~g,.!t'l
~~0.2<>: c:
~.20.1 <J) ~
.Q
«
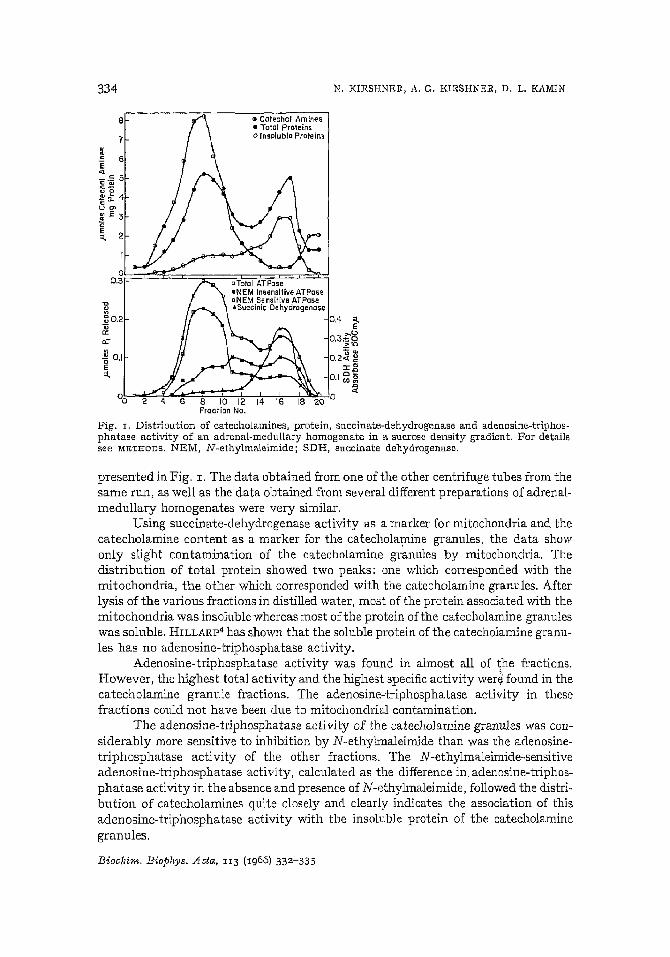
Fig. J. Distribution of catecholamines, protein, succinate-dehydrogenase and adenosine-triphosphatase activity of an adrenal-medullary homogenate in a sucrose density gradient. For detailssee METHODS. NEM, N-ethylmaleimide; SDH, succinate dehydrogenase.
presented in Fig. 1. The data obtained from one of the other centrifuge tubes from thesame run, as well as the data obtained from several different preparations of adrenalmedullary homogenates were very similar.
Using succinate-dehydrogenase activity as a marker for mitochondria and thecatecholamine content as a marker for the catecholamine granules, the data showonly slight contamination of the catecholamine granules by mitochondria. Thedistribution of total protein showed two peaks: one which corresponded with themitochondria, the other which corresponded with the catecholamine granules. Afterlysis of the various fractions in distilled water, most of the protein associated with themitochondria was insoluble whereas most of the protein ofthe catecholamine granuleswas soluble. HILLARp 4 has shown that the soluble protein of the catecholamine granules has no adenosine-triphosphatase activity.
Adenosine-triphosphatase activity was found in almost all of the fractions.However, the highest total activity and the highest specific activity were found in thecatecholamine granule fractions. The adenosine-triphosphatase activity in thesefractions could not have been due to mitochondrial contamination.
The adenosine-triphosphatase activity of the catecholamine granules was considerably more sensitive to inhibition by N-ethylmaleirnide than was the adenosinetriphosphatase activity of the other fractions. The N-ethylmaleimide-sensitiveadenosine-triphosphatase activity, calculated as the difference in adenosine-triphosphatase activity in the absence and presence of N-ethylmaleimide, followed the distribution of catecholamines quite closely and clearly indicates the association of thisadenosine-triphosphatase activity with the insoluble protein of the catecholaminegranules.
Biochim, Biophys, Acta, II3 (1966) 332-335

ATPASE ACTIVITY OF CATECHOLAMINE GRANULES 335
The possibility that the adenosine-triphosphatase activity of the catecholaminefraction might be due to ribosomes or lysosomes is remote. Little ribosomal contamination would be expected in the fractions and the RNA content of the variousfractions was quite low and could not be con-elated with the peaks of adenosine-triphosphatase activity. One cannot yet generalize about the density of lysosomes butin most tissues the lysosomes are less dense than mitochondria. In addition, most ofthe hydrolytic enzymes oflysosomes are solubilized upon lysis and only slight amountsof esterase activity are associated with the membranes.
The data obtained in these experiments confirm HILLARP'S report that thecatecholamine granules contain an adenosine-triphosphatase activity. AlthoughFORTIER et at.5 concluded that the adenosine-triphosphatase activity of their preparations was due to mitochondrial contamination, their data are not inconsistent withthose of HILLARP'S or ours. In fact, their data indicate that not all of the adenosinetriphosphatase activity in their granule fraction could have been due to mitochondrialcontamination. An additional factor that may explain the relatively low adenosinetriphosphatase activity of the granule fractions found by FORTIER et at.s is that theyused low concentrations of Ca2+, 2.5 mM, whereas HILLARP and we used 5 rrrMMgC12.
It was observed during the course of this work that low concentrations of Ca2+stimulated the adenosine-triphosphatase activity associated with the mitochondriato a greater extent than it did the adenosine-triphosphatase activity of the granulefractions.
ACKNOWLEDGEMENTS
This work was supported by U.S. Public Health Service Grant AM 05427.One of the authors (N.K.) was a career development awardee of the U.S. PublicHealth Service (K3-GM I5 I84).
REFERENCES
I A. CARLSSON, N.-A. HILLARP AND B. WALDECK, Acta Pliysiol, Scand. Suppl., 59 (1963) 215.2 N. KIRSHNER, ]. Bioi. Chem., 237 (1962) 2311.3 N. KIRSHNER, in G. B. KOELLE, W. W. DOUGLAS AND A. CARLSSON, Pharmacology oj Chot-
energic and Adrenergic Transmission, Pergamon Press, Oxford, 1966, p. 225.4 N.-A. I-IrLLARP, Acta Physiol, Scand., 43 (1958) 82.5 A. FORTIER, ]. LEDUC AND A. D'IoRIO, Rev. Can. BiD,.., 18 (1959) IIO.
6 P. HAGEN, Biochem, ]., 82 (1962) 25P.7 U. S. VON EULER AND U. HAMBERG, Science, IIO (1949) 561.8 ~. MOORE AND W. H. STEIN, J. Biol. Chem., 211 (1954) 907.9 H. I-I. TAUSSKY AND E. THORR, J. Biot. Chem., 202 (195)) 675.
10 C. H. WILLIAMS JR. AND H. KAMIN, .1. Bioi. Chem., 237 (1962) 587.II P. BANKS, Biochem, J., 95 (1965) 490.12 H. BLASCHKO, A. D. SMITH AND H. WINKLER, Biochem, J., 196 (1965) 49 P.13 P. HAGEN AND A. D'IoRIO, Cat/. J. Biochem., 43 (1965) 1633.
NOTE ADDED IN PROOF: After this manuscript was submitted, three other publications onthe ATPase activity of adrenal chromaffin granules have appeared. Banks" and Blaschke et al.ahave also found ATPase activity associated with the chromaffin granule fraction. Blaschke et al.I O
have also separated the ATPase activity of the chromaffin granules from several typical lysosomalenzyme activities by centrifugation through sucrose density g-radients similar to those used in ourlaboratory. However, Hagen and D'Eorio-", also using similar density gradients, have found only lowamounts of A'I'Pase activity in the chromaffin granule fraction and attribute this activity tomicrosomal or mitochondrial contamination. The explanation of these differences is not apparent.(Received November r Sth, 1965).
Biocbim. Biopliys. Acta, II3 (1966) 332-335





























