Embed Size (px)
Citation preview

RESEARCH ARTICLE
Multisensory Processing in Children With Autism: High-DensityElectrical Mapping of Auditory–Somatosensory Integration
Natalie Russo, John J. Foxe, Alice B. Brandwein, Ted Altschuler, Hilary Gomes, and Sophie Molholm
Successful integration of signals from the various sensory systems is crucial for normal sensory–perceptual functioning,allowing for the perception of coherent objects rather than a disconnected cluster of fragmented features. Several prominenttheories of autism suggest that automatic integration is impaired in this population, but there have been few empirical testsof this thesis. A standard electrophysiological metric of multisensory integration (MSI) was used to test the integrity ofauditory–somatosensory integration in children with autism (N 5 17, aged 6–16 years), compared to age- and IQ-matchedtypically developing (TD) children. High-density electrophysiology was recorded while participants were presented witheither auditory or somatosensory stimuli alone (unisensory conditions), or as a combined auditory–somatosensory stimulus(multisensory condition), in randomized order. Participants watched a silent movie during testing, ignoring concurrentstimulation. Significant differences between neural responses to the multisensory auditory–somatosensory stimulus and theunisensory stimuli (the sum of the responses to the auditory and somatosensory stimuli when presented alone) served asthe dependent measure. The data revealed group differences in the integration of auditory and somatosensory informationthat appeared at around 175ms, and were characterized by the presence of MSI for the TD but not the autism spectrumdisorder (ASD) children. Overall, MSI was less extensive in the ASD group. These findings are discussed within theframework of current knowledge of MSI in typical development as well as in relation to theories of ASD.
Keywords: autism spectrum disorders; electrophysiology; multisensory integration; auditory processing; somatosensoryprocessing; development
Introduction
Anecdotal reports from parents, clinicians and indivi-
duals with autism have lead to the impression that there
are significant sensory processing atypicalities in a large
percentage of individuals with an autism spectrum
disorder (ASD) [Cesaroni & Garber, 1991; Grandin,
1992; O’Neill & Jones, 1997; Williams, 1994]. In the
1970’s, sensory processing research was central to the
field of ASD, with researchers finding evidence for
atypical sensory modulation [e.g. Stroh & Buick, 1964],
a preferential reliance on proximal rather than distal
senses [Hermelin & O’Conner, 1970], a dominance of
somatosensory over auditory stimuli [Hermelin & O’Con-
ner, 1970], a tendency to be overselective in responding
to only one aspect of a multisensory object [e.g. Lovaas &
Schreibman, 1971; Lovaas, Schreibman, Koegal, & Rehm,
1971], and a tendency to benefit less from visual cues in a
visual-motor task than typically developing (TD) children
[Hermelin & O’Conner, 1970]. Despite these findings, the
research on sensory processing in ASD fell off in the
1980’s and 1990’s, to be replaced by research on higher-
order processes, such as executive function [e.g. Ozonoff
& Strayer, 1997], language [e.g. Tager-Flusberg, 1996], and
social difficulties [e.g. Klin et al., 1999; Tager-Flusberg &
Sullivan, 1994], that are also common among persons
with ASDs. However, a paradigm shift that once again
emphasizes basic sensory processing differences among
persons with ASD has emerged in recent years, as
evidenced by a resurgence of research utilizing ques-
tionnaire methods to describe sensory processing diffi-
culties [e.g. Crane, Goddard, & Pring, 2009; Lane, Young,
Baker, & Angley, 2010; Schoen, Miller, Brett-Green, &
Nielsen, 2009; Tomchek, 2007], and an emerging litera-
ture on unisensory detection and discrimination in ASD
[e.g. Bertone, Mottron, Jelenic, & Faubert, 2003, 2005;
Bonnel et al., 2003; O’Riordan, 2004; O’Riordan &
Plaisted, 2001; O’Riordan & Passetti, 2006]. This shift is
particularly evident in the impending changes in the
diagnosis of ASD expected in the DSM-V, in which social
INSAR Autism Research 3: 1–15, 2010 1
Received July 13, 2009; accepted for publication June 25, 2010
Published online in Wiley Online Library (wileyonlinelibrary.com)
DOI: 10.1002/aur.152
& 2010 International Society for Autism Research, Wiley Periodicals, Inc.
From the City College of New York, The Children’s Research Unit, Program in Cognitive Neuroscience, Departments of Psychology & Biology, New York
(N.R., J.J.F., T.A., H.G., S.M.,); Albert Einstein College of Medicine, Children’s Evaluation and Rehabilitation Center (CERC); Departments of Pediatrics
and Neuroscience, Bronx, New York (N.R., J.J.F., A.B.B., T.A., S.M.,); Queens College of the City University of New York, Flushing, New York (J.J.F., A.B.B.,
H.G.)
Address for correspondence and reprints: Sophie Molholm, Albert Einstein College of Medicine, Children’s Evaluation and Rehabilitation Center
(CERC), Departments of Pediatrics and Neuroscience, Jack & Pearl Resnick Campus, 1300 Morris Park Avenue, Van Etten 1st floor, C-wing, Bronx, NY,
10461. E-mail: [email protected]
Grant sponsor: NIH; Grant number: MH 085322; Grant sponsors: Wallace Research Foundation; Cure Autism Now; Fondation du Quebec de Recherche
sur la Societe et la Culture; Canadian Institute of Health Research.

and communication difficulties are being combined into
one diagnostic criterion, thus placing equal weight on
both the negative (language, socialization, and commu-
nication) and positive symptoms (repetitive behaviors,
restricted interests, and sensory issues) of autism.
One suggestion that has gained prominence in the
autism community, but that has only recently become
the subject of empirical investigation, is that a problem
with the integration of sensory information may underlie
some of the symptoms observed in autism, including the
observed sensory hyper- and hypo-sensitivities. Despite
renewed interest in sensory issues and the prominence of
so-called sensory integration treatment centers and books
written on this topic [e.g. Ayers, 1994], surprisingly little
empirical work has been conducted to actually test the
integrity of multisensory integration (MSI) in autism.
MSI in Typical Development
In adults, redundant or mutually informative multi-
sensory inputs tend to be effortlessly integrated, and
often lead to faster and better performance on discrimi-
nation and detection tasks relative to performance on
their unisensory counterparts [e.g. Frens & Van Opstal,
1995; Molholm et al., 2002; Molholm, Ritter, Javitt, &
Foxe, 2004; Murray et al., 2005; Perrott, Saberi, Brown, &
Strybel, 1990; Spence & Driver, 1997; Stein & Meredith,
1990; Zahn, Abel, & Dell’Osso, 1978]. The same is the
case for more complex perceptual-cognitive processes,
with faster identification of objects when multisensory
inputs are provided and better speech recognition when
the speaker’s articulations can be seen compared to when
they cannot [Driver, 1996; Molholm et al., 2004; Ross,
St-Amour, Leavitt, Javitt, & Foxe, 2007]. The ability to
recognize lawful relationships between multisensory
inputs is seen from a very early age, with infants able to
recognize that audiovisual stimuli are temporally asyn-
chronous as early as 4 months of age [Lewkowicz, 2002].
Natural ‘‘sensory deprivation’’ in humans in cases of early
blindness [Hotting, Rosler, & Roder, 2004; Putzar,
Goerendt, Lange, Rosler, & Roder, 2007], and controlled
sensory deprivation studies in animals [Carriere et al.,
2007; Wallace, Perrault, Hairston, & Stein, 2004] reveal
that, as might be expected, post-natal interaction with
the environment plays a significant role in the develop-
ment of MSI. Developmental studies also reveal that for
some types of information, MSI processes tune-up to
optimal levels over the first decade or two of life
[Brandwein et al., accepted; Gori, Del Viva, Sandini, &
Burr, 2008; Nardini, Jones, Bedford, & Braddick, 2008].
MSI in ASDs
While it has been speculated that multisensory proces-
sing is impaired in autism, there has been little empirical
testing of this notion [see Foxe & Molholm, 2009]. What
studies there are have all examined interactions between
auditory and visual sensory inputs, and these studies
have yielded equivocal results. For example in one study,
temporal asynchrony judgments were impaired com-
pared to matched controls for linguistic stimuli but not
for non-linguistic stimuli [Bebko, Weiss, Demark, &
Gomez, 2006]. In another, Smith and Bennetto [2007]
found that children and adults with ASD were less
susceptible to the McGurk illusion, in which the percept
of a speech sound is changed by the simultaneous
presentation of an incongruous visual speech articula-
tion. This suggests that these auditory and visual speech
inputs were not integrated in a typical manner in ASD, a
finding receiving additional support from work by de
Gelder, Vroomen, and van der Heide [1991] who noted
that in spite of normal lip-reading ability, individuals
with ASD showed less influence of visual speech on
auditory speech perception than mental age matched TD
individuals. In contrast, van der Smagt, van Engeland,
and Kemner [2007], found that children with ASD were
just as susceptible to an audiovisual illusion as were TDs.
For both groups, hearing multiple sounds resulted in the
illusory perception of multiple flashes when only a single
flash was presented [the beep-flash illusion, see Shams,
Kamitani, & Shimojo, 2000, 2002].
Cross-sensory priming paradigms, in which a visual
stimulus precedes an auditory stimulus and its effects on
auditory processing are examined, have demonstrated
intact visual modulation of the auditory electrophysio-
logical response among persons with an ASD [Magnee, de
Gelder, van Engalend, & Kemner, 2008; Magnee, Oranje,
van Engeland, Kahns, & Kemner, 2009]. In these studies a
visual stimulus was presented 350–500 ms before the
onset of an auditory stimulus (in one case the visual
stimulus remained on for the duration of the auditory
stimulus and in the other it did not), and suppression of
the auditory-N1 response was observed. While these data
have been interpreted as evidence for intact early multi-
sensory processing in ASD, Vroomen and Stekelenburg
[2009] have convincingly demonstrated that this sort of
cross-sensory N1 suppression is due to the predictive
properties of the preceding visual stimulus on onset of
the upcoming auditory stimulus, and not multisensory
processing per say. Hence, preservation of such effects in
ASD cannot be taken as evidence for intact multisensory
processing.
The literature on multisensory processing in ASD thus
far has largely been confined to studies that have
required behavioral responses to assess multisensory
function [but see Magnee et al., 2008, 2009]. While
behavioral measures are certainly valuable, a disadvan-
tage of this approach is that it rests on the assumption
that observed differences largely reflect multisensory
processing, whereas behavioral differences could well be
due to differing interpretation of task instruction or
2 Russo et al./Auditory–somatosensory integration in ASD INSAR

differences in how attention is allocated (e.g. superior
attentional allocation for one group over the other). In
either of these cases, the implications are clearly
different. In addition, observed similarities in behavior
do not necessarily mean that the underlying brain
processes are the same. Electrophysiology provides a
metric of multisensory processing at multiple stages of
the processing hierarchy, and can be generated in the
absence of attentional demands through the use of
‘‘passive’’ paradigms in which there is no task. As a
result, the integrity of MSI can be assessed without
confounding group differences in attentional allocation
or task comprehension.
Electrophysiology of MSI in Typical Development
Both passive and active paradigms have been used to
assess the electrophysiology of auditory–somatosensory
integration in TD adults and children, with consistent
findings that MSI occurs at multiple processing stages
starting early on (circa 50 ms) in cortical areas that were
traditionally considered to be unisensory [e.g. Foxe et al.,
2000; Lutkenhoner, Lammertmann, Simoes, & Hari,
2002]. In one of the first published electrophysiological
studies of MSI in children, Brett-Green, Miller, Gavin, and
Davies [2008] examined multisensory processing in
children between the ages of 6 and 13 years. Using
median nerve stimulation and tones presented alone or
together, the authors found three time periods during
which multisensory interactions occurred: 60–80 ms over
central scalp regions of the hemisphere contralateral to
the stimulated side, 110–150 ms in the hemisphere
ipsilateral to the side of the stimulation over central-
parietal areas, and 180–220 ms over central regions
bilaterally. These latencies, though slightly later than
those found in adults [e.g. Foxe et al., 2002; Murray et al.,
2005; Sperdin, Cappe, Foxe, & Murray, 2009], support the
notion that children, like adults, integrate multisensory
information during both early and later sensory–percep-
tual time frames.
In the present study, high-density electrophysiology
was used to test the integrity of multisensory processing
of auditory and somatosensory information among
persons with an ASD. Auditory and somatosensory
stimuli were presented in randomized order, either alone
or simultaneously, in a passive paradigm in which
participants engaged in an unrelated activity (watching
a silent video). MSI was indexed by comparing electro-
physiological responses to the stimuli when presented
alone vs. when presented together [e.g. Berman, 1961;
Brett-Green et al., 2008; Foxe et al., 2000, 2002; Giard &
Peronnet, 1999; Molholm et al., 2002; Murray et al.,
2005; Talsma & Woldorff, 2005; Teder-Salejarvi, McDonald,
DiRusso, & Hillyard, 2002]. This commonly used index of
MSI entails summing the responses to the auditory and
somatosensory stimuli when presented alone and com-
paring this ‘‘sum’’ waveform to the response to the
auditory and somatosensory stimuli when presented
together. Because electrical fields sum linearly at the
scalp, any differences between the two responses can
then be attributed to the two inputs having interacted
when they were presented together.1 Our working
hypothesis was that this electrophysiological metric
would reveal differences in multisensory processing
between individuals with ASD and TD, age- and
IQ-matched children. Specifically, we predicted a reduc-
tion or absence of MSI in ASD at earlier time points,
which we assume to reflect more automatic integrative
processes, and a corresponding increase in MSI at later
time points (4200 ms) reflecting compensatory and less
automatic processing.
MethodsParticipants
Seventeen children with ASD and 17 TD children
between the ages of 6 and 16 years participated in this
study (see Table 1). In accordance with the Declaration of
Helsinki, the parents of all the participants provided a
written informed consent, and when appropriate, chil-
dren provided a written assent. All the procedures and
consent forms were approved by the Institutional Review
Board of the City College of New York.
Exclusionary criteria for both groups included uncor-
rected vision problems, a history of seizures, and the use
of psychotropic medication. TD children were excluded if
they had a history of educational, attentional, psychiatric,
or developmental difficulties as assessed by a history
questionnaire, and were also excluded if their parents
endorsed six items of inattention or hyperactivity on a
Table 1. Participant Characteristics for the Autism SpectrumDisorder (ASD) and Typically Developing (TD) Groups
Age Verbal IQ Performance IQ Full-scale IQ
TD 10.48(2.92) 110.3 (11.83) 106.53 (13.35) 108.4 (12.65)
ASD 10.36 (2.71) 102.52(18.1) 105.06 (14.24) 104.7 (14.58)
P-values 0.9 0.15 0.76 0.3
Values within parenthesis represent SD.
1The assumption of linearity no longer holds for task-related processes
that would be shared for the two unisensory inputs in an active paradigm,
such as when each would result in the recruitment of motor cortex to
make a button press response. This is because this process would be
doubly represented in the summed response and only represented once in
the multisensory response. However, in the present study, we are dealing
with a passive paradigm and focusing mainly on the sensory processing
time frame, and hence the assumption of linearity is expected to hold,
with its violation representing the occurrence of multisensory interac-
tions in the brain.
INSAR Russo et al./Auditory–somatosensory integration in ASD 3

DSM-IV behavioral checklist of attention deficit disorder
(with and without hyperactivity). Both forms were filled
out by parents on the first day of testing. The parents of
children with ASD were asked to refrain from giving their
children stimulant medication (n 5 2) before the testing
session.
Diagnoses for 15 of the 17 children with ASD were
made using both the Autism Diagnostic Interview-R
[ADI-R; Lord, Rutter, & LeCouter, 1994] and the Autism
Diagnostic Observation Schedule [ADOS-G; Lord, Rutter,
DiLavore, & Risi, 1999] by a trained administrator. For the
remaining two, who were diagnosed with Pervasive
Developmental Disorder, Not Otherwise Specified (PDD-
NOS), ADOS, and ADI, data were not available and
diagnosis was made on the basis of the diagnostic criteria
outlined in the DSM-IV-TR [APA, 2000]. Of the 17 ASD
participants, seven had autism, eight had Asperger’s
disorder, and two had a PDD-NOS. The children with
ASD and the TD children were matched on a one-to-one
basis, and within one standard deviation on Full Scale
and Performance IQ’s (FSIQ and PIQ) as measured by the
Wechsler Abbreviated Test of Intelligence [Wechsler,
1999], as well as on chronological age. The two groups
did not differ significantly on any of the matching
measures nor on Verbal IQ (see Table 1). All of the
children had IQs in the average to above average range
with the exception of one TD and one ASD child with
IQ’s in the borderline range.
Stimuli and Procedure
Testing was conducted over 2 days. In a first session,
participants completed the diagnostic and psychometric
evaluation. Event-Related Potential (ERP) recordings were
conducted on a second visit in a sound attenuated, dimly
lit electrically shielded room. Participants were seated in
a comfortable chair and watched a film of their choice
without sound during the testing protocol, which was
presented in three blocks of 10 min each.
Somatosensory stimulation. Somatosensory stimu-lation consisted of a 128-Hz vibrotactile stimulationpresented to the child’s right hand for 30ms. Thestimulation was delivered through a 2-layer Transducermanufactured by Piezo Systems, Inc, which was driven bythe soundcard of a PC through an audio cable. Thestimulator was held with gauze between the child’sthumb, index, and middle finger, and the child’s handwas covered by a black cloth, as seeing somatosensorystimulation can affect the somatosensory response [e.g.Taylor-Clarke, Kennett, & Haggard, 2002].
Auditory stimulation. Auditory stimulation consistedof a 1000Hz tone presented for 30ms through a speakerplaced on the child’s right side. The somatosensory andauditory stimuli could either be presented alone (unisensoryconditions) or simultaneously (multisensory condition) for
a total of three stimulus conditions: auditory (A: 300 trials),somatosensory (S: 300 trials) and multisensory (AS: 300trials), totaling 900 trials. The conditions were presented ina randomized order, with a randomized square-wavedistribution of Stimulus Onset Asynchrony’s (SOAs)ranging from 700 to 3,000ms. This large variance in SOAis an effective way to circumvent issues with anticipatoryslow-wave activity [see Teder-Salejarvi et al., 2002].
Measures and analyses. Sixty-eight-channel scalp EEGwas recorded, amplified, and digitized at 512 Hz usingBioSemi Systems Active Amplifiers. BioSemi uses twoelectrodes—the Common Mode Sense (CMS), which isactively recorded, and the Driven Right Leg (DRL), apassive electrode—that together form a feedback loopthat represent the reference. The acquisition of the dataoccurs referenced to the CMS-DRL ground which drivesthe average potential of the participant (i.e. the commonmode voltage) as close as possible to the AC referencevoltage of the Analog-to-Digital box (for a description ofthe BioSemi active electrode system referencing andgrounding conventions, visit www.biosemi.com/faq/cms&drl.htm).
The continuous raw EEG data were first visually
inspected to ensure sufficient data quality across our
subject population without regard for group membership
(TD or ASD). Based on this initial inspection and
preliminary exploratory analyses using preset artifact
rejection criteria, it was determined that 160 usable trials
per condition could reliably be extracted for the vast
majority of subjects and this number was then preset as
our target trial count. This strategy was motivated by a
wish to equate the number of trials sampled from
participants from both populations. Specifically, 160
trials of each of the three conditions, distributed evenly
across the three stimulation blocks (55 trials from each
block), were selected by hand from the raw EEG to be
included in the individual subject averages, with the
restriction that the signal did not exceed the artifact
rejection criterion of plus or minus 130 microvolts. Note
that in almost all cases, there were considerably more
than 160 trials that met these criteria. An exception was
made in the case of just one ASD participant for whom
we could only identify 80 trials across each of the
stimulation types that met these criteria. In turn, we
sampled just 80 trials per condition from his matched
control participant. It is important to point out that
during the hand selection procedure, there was no
possibility that the evoked brain responses could be
visualized in the raw EEG and the experimenter was blind
as to group membership. Accordingly trial selection
proceeded in an unbiased manner. A second criterion
was applied to selection of trials containing somatosen-
sory stimulation. Trials were only chosen for inclusion in
averages for those conditions where the presence of a
prominent and typical stimulus-artifact from the soma-
tosensory stimulator could be visualized in the raw EEG.
4 Russo et al./Auditory–somatosensory integration in ASD INSAR

The latter was done as an assurance that somatosensory
stimulation had definitively occurred, which could be
verified on greater than 75% of trials. This telltale
stimulus-artifact results from skin conduction of the
electrical current of the stimulator to the electrodes on
the scalp. We took this cautious approach because the
lack of such a stimulus-artifact could potentially indicate
that the stimulator had not made full contact on those
trials where it was not fully evident, and we wanted to
protect against any possibility that trials without appro-
priate stimulation might be included in the averages,
although this was likely the case on only a very small
minority of trials.
For post-processing, a band-pass filter of 2.4 Hz at 12db/
octave–30 Hz at 12 db/octave was applied to the contin-
uous data. High-pass filtering was performed to remove
any ongoing slow-wave activity remaining in the signal,
which would otherwise be doubly represented in the
summed unisensory response when assessing MSIs [see
Molholm et al., 2002; Teder-Salejarvii et al., 2002]. Low-
pass filtering served to remove high-frequency artifact
that resulted from the somatosensory stimulator. The
EEG data were epoched into 600 ms segments that
included the 100-ms preceding stimulus onset, artifact
rejected at7130 mV, and averaged based on stimulation
type using Brain Electric Source Analysis version
(www.BESA.de). The data were re-referenced to the
average reference for all analyses and figures. For analysis
of MSIs, the auditory-alone and somatosensory-alone
responses were summed for comparison with the multi-
sensory condition.
Data Analysis
Analysis of the data was guided by previous studies that
have similarly used electrophysiology to assess MSI of
simple auditory and somatosensory stimuli [e.g. Foxe
et al., 2000]. These studies have shown a series of
multisensory modulations that fall on the major peaks
of the auditory–somatosensory response over the first
200 ms of processing. To constrain our analyses, we
therefore used the average of the grand-mean auditory–
somatosensory responses from each of the ASD and TD
groups to define the peaks of the multisensory response,
and constrained our planned analyses to windows
around these peaks. The electrodes tested were selected
based on the corresponding scalp distribution of the
responses at each of the latencies. This technique allowed
us to predefine time windows and scalp regions of
analysis, and to do so in a way that did not make
reference to the dependent measure. Significant interac-
tions with the factor of group were examined by
conducting univariate Repeated Measures ANOVA’s.
The results of all the ANOVA’s are based on Green-
house–Geiser corrections.
Exploratory Analysis
In addition to the above conservative approach to data
analysis, exploratory ‘‘cluster plot’’ analyses were also
performed to fully describe the data. In this approach,
t-tests are performed for each data point for each
electrode and significance values are plotted for each of
these points and displayed in a cluster plot. To reduce the
probability of Type-1 errors, significant differences are
only considered if a criterion of Po0.05 is achieved for at
least 10 consecutive time points [Guthrie & Buchwald,
1991]. It should be pointed out that this criterion would
exclude any effects that did not last for at least 18 ms,
given our digitization rate of 512 Hz. This is a suitable
alternative to Bonferroni correction for multiple compar-
isons, which would increase the likelihood of Type II
errors through overcompensation for Type I errors. This
analysis has the advantage of providing a snapshot of all
significant differences within a single frame, and the
potential to reveal unpredicted effects that can then be
used as a hypothesis generation tool for future studies.
Using this approach, MSIs for each of the participant
groups were examined by comparing the multisensory
and the sum response. Using this same approach, each of
the auditory and somatosensory unisensory responses
were compared across the groups to probe for the
possibility of group differences in basic sensory proces-
sing.
ResultsUnisensory Responses
The electrophysiological responses to the auditory-alone
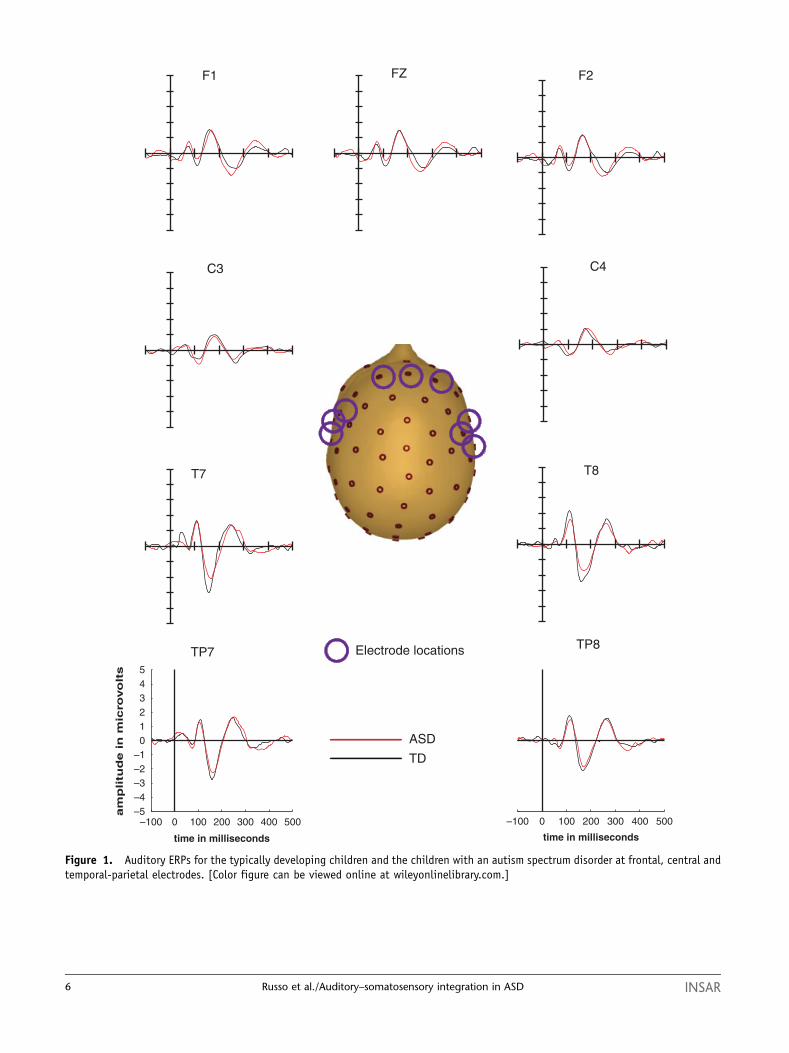
stimuli (Fig. 1) were highly similar in morphology
between the two participant groups. The scalp distribu-
tion and timing of the auditory response for both groups,
illustrated in Figure 1, represents an amalgam of typical
auditory-evoked responses for younger and older chil-
dren, with major responses both fronto-centrally and at
lateral temporal scalp regions [Ceponiene, Rinne &
Naatanen, 2002]. This is to be expected from our
relatively broad age sample. While the group responses
were highly similar, starting at 100 ms the ASD response
tended to be smaller in amplitude. This diminution of
the response can be clearly seen at the response peaks,
over fronto-central scalp at 100 ms and over bilateral
temporal scalp regions around 150 ms. The electrophy-
siological responses to the somatosensory-alone stimuli
(Fig. 2) also showed a similar morphology across the
groups, but here too responses appeared to be larger for
the TD group compared to the ASD group, starting at
about 70 ms. Somatosensory responses were the greatest
contralateral to the side of stimulation, over left central
scalp areas, with positive-going peaks at 53 and 110 ms.
This latter peak was more pronounced for the TD than
INSAR Russo et al./Auditory–somatosensory integration in ASD 5
TP7TP8
T7 T8
–100 0 100 200 300 400 500–5–4–3–2–1
012345
time in milliseconds
am
plitu
de in
mic
rovo
lts
–100 0 100 200 300 400 500
time in milliseconds
ASD
TD
F1 FZ F2
C4C3
Electrode locations
Figure 1. Auditory ERPs for the typically developing children and the children with an autism spectrum disorder at frontal, central andtemporal-parietal electrodes. [Color figure can be viewed online at wileyonlinelibrary.com.]
6 Russo et al./Auditory–somatosensory integration in ASD INSAR
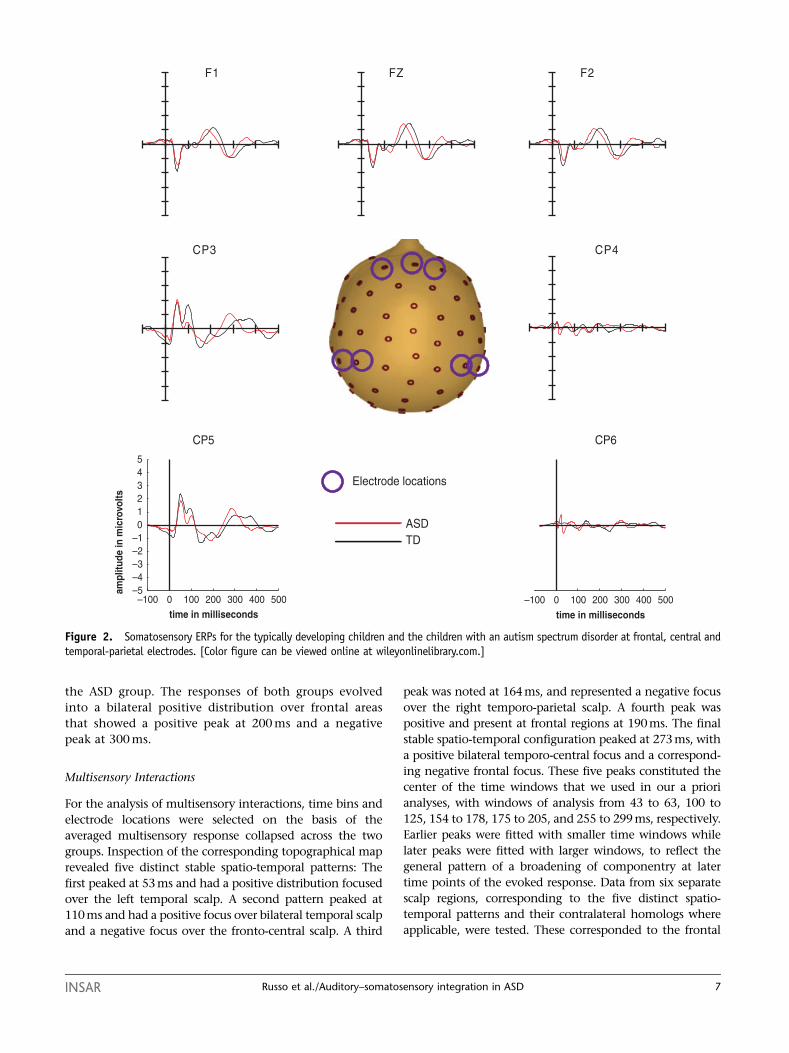
the ASD group. The responses of both groups evolved
into a bilateral positive distribution over frontal areas
that showed a positive peak at 200 ms and a negative
peak at 300 ms.
Multisensory Interactions
For the analysis of multisensory interactions, time bins and
electrode locations were selected on the basis of the
averaged multisensory response collapsed across the two
groups. Inspection of the corresponding topographical map
revealed five distinct stable spatio-temporal patterns: The
first peaked at 53ms and had a positive distribution focused
over the left temporal scalp. A second pattern peaked at
110ms and had a positive focus over bilateral temporal scalp
and a negative focus over the fronto-central scalp. A third
peak was noted at 164ms, and represented a negative focus
over the right temporo-parietal scalp. A fourth peak was
positive and present at frontal regions at 190ms. The final
stable spatio-temporal configuration peaked at 273ms, with
a positive bilateral temporo-central focus and a correspond-
ing negative frontal focus. These five peaks constituted the
center of the time windows that we used in our a priori
analyses, with windows of analysis from 43 to 63, 100 to
125, 154 to 178, 175 to 205, and 255 to 299ms, respectively.
Earlier peaks were fitted with smaller time windows while
later peaks were fitted with larger windows, to reflect the
general pattern of a broadening of componentry at later
time points of the evoked response. Data from six separate
scalp regions, corresponding to the five distinct spatio-
temporal patterns and their contralateral homologs where
applicable, were tested. These corresponded to the frontal
2FZF1F
4PC3PC
–100 0 100 200 300 400 500–5–4–3–2–1012345
time in milliseconds
ampl
itude
in m
icro
volts
CP6CP5
–100 0 100 200 300 400 500
time in milliseconds
ASDTD
Electrode locations
Figure 2. Somatosensory ERPs for the typically developing children and the children with an autism spectrum disorder at frontal, central andtemporal-parietal electrodes. [Color figure can be viewed online at wileyonlinelibrary.com.]
INSAR Russo et al./Auditory–somatosensory integration in ASD 7

(F1, Fz, and F2), fronto-central (FC1, FCz, and FC2), left
central (T7, C5, and C3), right central (T8, C6, and C4), left
temporal (TP7, CP5, and CP3), and the right temporal (TP8,
CP6, and CP4) regions. The regions tested for each of the
specific time windows were as follows: left and right
temporal regions (TP7, CP5, CP3, T8, CP6, and CP4) were
tested for the 43- to 63-ms window, left and right temporal
regions (TP7, CP5, CP3, T8, CP6, and CP4), and left and
right central regions (T7, C5, C3, T8, C6, and C4) were
tested for the 100- to 125-ms time window, left and right
temporal (TP7, CP5, CP3, TP8, CP6, and CP4) were tested for
the 154- to 178-ms time window, frontal central (FC1, FCz,
and FC2) and frontal regions (F1, Fz, and F2) were tested for
the 175- to 205-ms time window, and finally, frontal central
(FC1, FCz, and FC2), left and right temporal (TP7, CP5, CP3,
T8, CP6, and CP4), and left and right central regions (T7,
C5, C3, T8, C6, and C4) and were tested for the 255- to 299-
ms window. The multisensory and ‘‘sum’’ ERPs were
compared using separate repeated measures ANOVA’s for
each time bin of analysis on the average referenced data,
with the between-subjects factor of group and the within-
subjects factors of condition (summed vs. multisensory) and
the corresponding region factors. Waveforms from repre-
sentative electrodes are presented in Figures 3 and 4 for
the TD and ASD groups respectively.
The analysis of data from 43 to 63 ms yielded no main
effect of condition or any significant interactions. In the
100- to 125-ms window, there was a main effect of
condition (F (1, 32) 5 6.121, P 5 0.019), providing
evidence of MSI. Neither group nor region interacted
significantly with condition (Ps40.53). In the 154- to
178-ms time window, there was a condition by region
interaction (F (1, 32) 5 9.91, P 5 0.004). This interaction
showed that the difference between sum and multi-
sensory responses was greater at the left (F (1, 32) 5 5.97,
P 5 0.02) than at the right temporal regions (F (1, 32) 5 4.2,
P 5 0.049). In the 175- to 205-ms time window, there was a
main effect of condition (F (1, 32) 5 4.727, P 5 0.037) and a
condition by group interaction that approached significance
(F (1, 32) 5 3.43, P 5 0.073), which was found in a
follow-up analysis to be related to the presence of significant
differences between summed and multisensory responses
for the TD group (F (1, 32) 5 8.11, P 5 0.008) but not the
ASD group (F (1, 32) 5 .051, P 5 0.822). Finally, in the 255-
to 299-ms time window, there was a condition by group
interaction (F (1, 32) 5 4.94, P 5 0.033), which in a follow-
up analysis was found to be related to the presence of
significant differences between the summed and multi-
sensory responses for the TD (F (1, 32) 5 5.3, P 5 0.028) but
not the ASD group (F(1, 32) 5 0.717, P 5 0.4).
Exploratory Analyses
Unisensory responses. In the exploratory analysis ofbetween-group differences in the processing ofunisensory stimuli, the statistical cluster plots
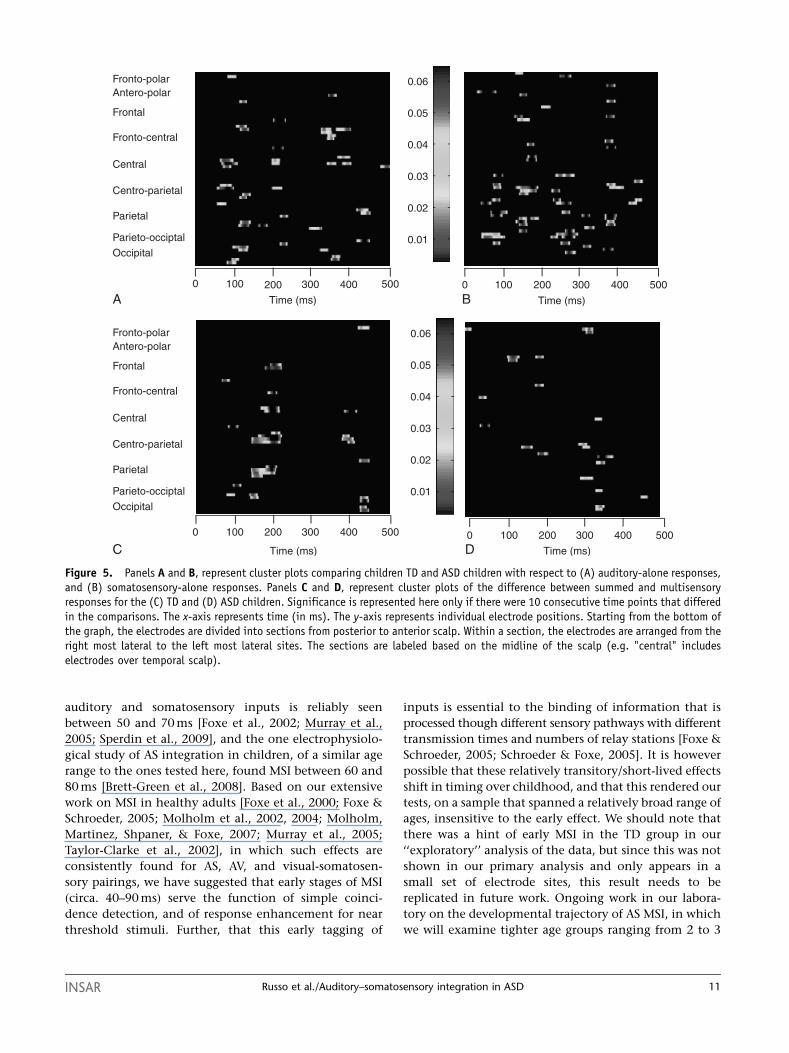
comparing the auditory responses revealed four timeperiods of significant difference between TD and ASDchildren (see panel A of Fig. 5). A first early difference wasapparent around 85 ms over central regions,characterized as a slightly larger P1 response for thegroup of children with ASD, a second difference around110 ms over parietal and frontal scalp regionsrepresenting a smaller N1 response in the ASD group, athird around 210 ms over more central scalp regions,which seemed to represent an N2 response that wasshifted earlier in time for the children with ASD, and afourth late difference around 350 ms over fronto-centralscalp regions, which reflected a larger P3 response for thechildren with ASD. Turning now to the between-groupdifferences in the somatosensory response (see panel B ofFig. 5), there were differences at about 70 and 100 ms,reflecting the smaller positive-going responses at theselatencies in the ASD group over the left centro-parietalregions. A difference was also seen at 150 ms due to asmaller negative going response in the ASD group overthe left centro-parietal scalp. Other differences werenoted at around 280 ms over the centro-parietal andparietal regions, with the somatosensory response in thistime frame appearing to occur earlier for the ASD thanthe TD group. Finally, late differences were noted againaround 380 ms over frontal regions, which reflected agreater response in the ASD compared to the TD group(see panel B of Fig. 5).
Multisensory responses. Exploratory analysis of MSIsusing the statistical cluster plots painted astraightforward picture of the MSIs in the ASD and TDgroups (see Panels C and D of Fig. 5). Specifically, theplots revealed a pattern of MSI from about 120–200 msthat was present in the TD group but not in the ASDgroup. This timing encompassed the initial three MSIeffects established in our a priori analysis. For the ASDgroup, a clear pattern of MSI only emerged at about300 ms (from �325 to 375 ms), a time frame that was notincluded in our a priori analysis. In the TD group, therewas also evidence for a second MSI effect at about 400 ms.Lowering the criterion to a less restrictive one of fivesuccessive significant points failed to reveal anyadditional clear patterns in the ASD group beyond thatat 300 ms, but did serve to solidify the effects at �120 and400 ms in the TD group, and also showed short-livedeffects that fell into the 250- to 300-ms range that wassignificant for this group in the ANOVA. There may evenbe a hint of an earlier effect at about 64 ms in the TDgroup that would bear replication in a follow-up study.
Discussion
The notion that fundamental differences in MSI are core
to ASD has been at the forefront of a number of
influential theories of the disorder since its original
description by Kanner in the 1940’s. In the present
study, the integrity of MSI in children with ASD was
8 Russo et al./Auditory–somatosensory integration in ASD INSAR
interrogated with scalp-recorded electrophysiology using
a passive auditory–somatosensory stimulation paradigm.
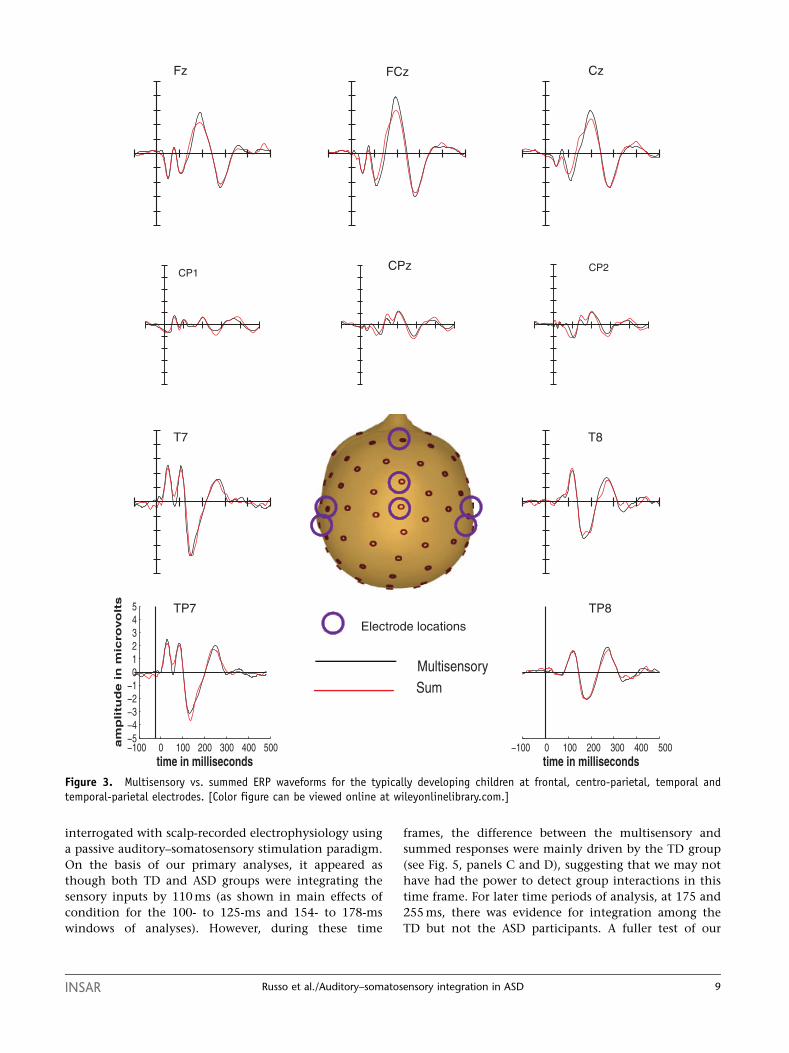
On the basis of our primary analyses, it appeared as
though both TD and ASD groups were integrating the
sensory inputs by 110 ms (as shown in main effects of
condition for the 100- to 125-ms and 154- to 178-ms
windows of analyses). However, during these time
frames, the difference between the multisensory and
summed responses were mainly driven by the TD group
(see Fig. 5, panels C and D), suggesting that we may not
have had the power to detect group interactions in this
time frame. For later time periods of analysis, at 175 and
255 ms, there was evidence for integration among the
TD but not the ASD participants. A fuller test of our
Figure 3. Multisensory vs. summed ERP waveforms for the typically developing children at frontal, centro-parietal, temporal andtemporal-parietal electrodes. [Color figure can be viewed online at wileyonlinelibrary.com.]
INSAR Russo et al./Auditory–somatosensory integration in ASD 9
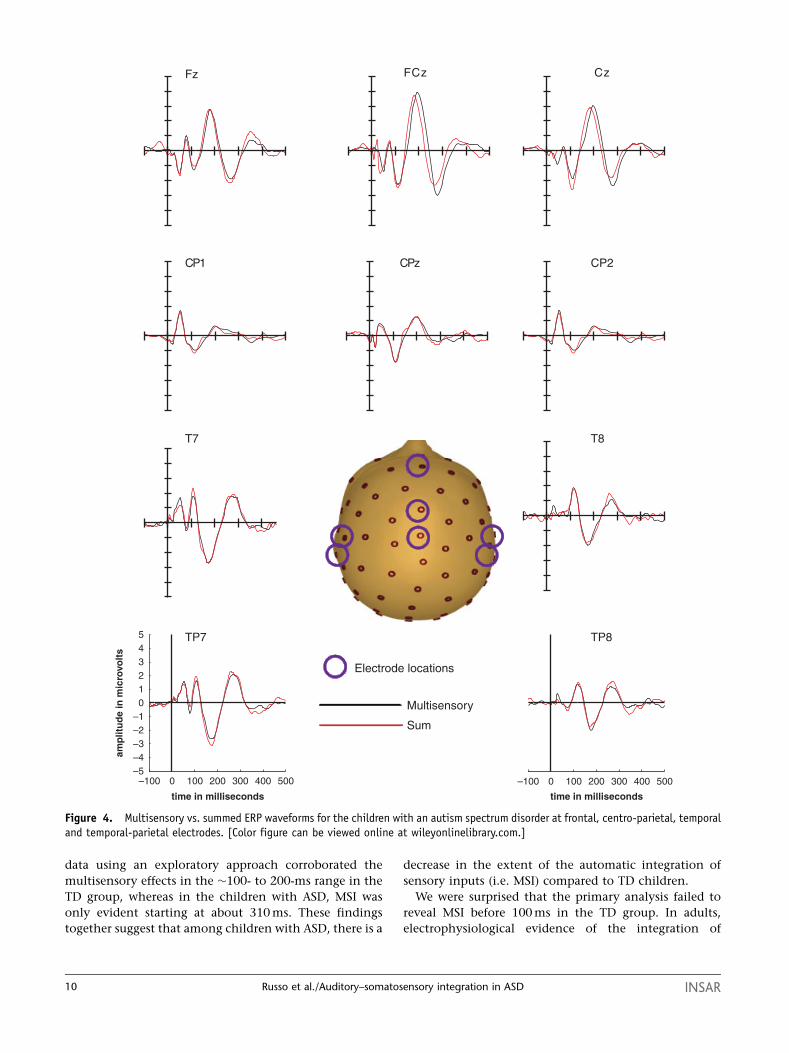
data using an exploratory approach corroborated the
multisensory effects in the �100- to 200-ms range in the
TD group, whereas in the children with ASD, MSI was
only evident starting at about 310 ms. These findings
together suggest that among children with ASD, there is a
decrease in the extent of the automatic integration of
sensory inputs (i.e. MSI) compared to TD children.
We were surprised that the primary analysis failed to
reveal MSI before 100 ms in the TD group. In adults,
electrophysiological evidence of the integration of
zCzCF
T7
Multisensory
Sum
–100 0 100 200 300 400 500–5–4–3–2–1012345
time in milliseconds
amp
litu
de
in m
icro
volt
s
–100 0 100 200 300 400 500
time in milliseconds
Electrode locations
Fz
TP8TP7
CP2zPC1PC
T8
Figure 4. Multisensory vs. summed ERP waveforms for the children with an autism spectrum disorder at frontal, centro-parietal, temporaland temporal-parietal electrodes. [Color figure can be viewed online at wileyonlinelibrary.com.]
10 Russo et al./Auditory–somatosensory integration in ASD INSAR
auditory and somatosensory inputs is reliably seen
between 50 and 70 ms [Foxe et al., 2002; Murray et al.,
2005; Sperdin et al., 2009], and the one electrophysiolo-
gical study of AS integration in children, of a similar age
range to the ones tested here, found MSI between 60 and
80 ms [Brett-Green et al., 2008]. Based on our extensive
work on MSI in healthy adults [Foxe et al., 2000; Foxe &
Schroeder, 2005; Molholm et al., 2002, 2004; Molholm,
Martinez, Shpaner, & Foxe, 2007; Murray et al., 2005;
Taylor-Clarke et al., 2002], in which such effects are
consistently found for AS, AV, and visual-somatosen-
sory pairings, we have suggested that early stages of MSI
(circa. 40–90 ms) serve the function of simple coinci-
dence detection, and of response enhancement for near
threshold stimuli. Further, that this early tagging of
inputs is essential to the binding of information that is
processed though different sensory pathways with different
transmission times and numbers of relay stations [Foxe &
Schroeder, 2005; Schroeder & Foxe, 2005]. It is however
possible that these relatively transitory/short-lived effects
shift in timing over childhood, and that this rendered our
tests, on a sample that spanned a relatively broad range of
ages, insensitive to the early effect. We should note that
there was a hint of early MSI in the TD group in our
‘‘exploratory’’ analysis of the data, but since this was not
shown in our primary analysis and only appears in a
small set of electrode sites, this result needs to be
replicated in future work. Ongoing work in our labora-
tory on the developmental trajectory of AS MSI, in which
we will examine tighter age groups ranging from 2 to 3
Fronto-polarAntero-polar
Frontal
Fronto-central
Central
Centro-parietal
Parietal
Parieto-occiptal
Occipital
0.06
0.05
0.04
0.03
0.02
0.01
0.06
0.05
0.04
0.03
0.02
0.01
Fronto-polarAntero-polar
Frontal
Fronto-central
Central
Centro-parietal
Parietal
Parieto-occiptal
Occipital
Time (ms)
0 100 200 300 500400
Time (ms)
0 100 200 300 500400
Time (ms)
0 100 200 300 500400
Time (ms)
0 100 200 300 500400
A B
C D
Figure 5. Panels A and B, represent cluster plots comparing children TD and ASD children with respect to (A) auditory-alone responses,and (B) somatosensory-alone responses. Panels C and D, represent cluster plots of the difference between summed and multisensoryresponses for the (C) TD and (D) ASD children. Significance is represented here only if there were 10 consecutive time points that differedin the comparisons. The x-axis represents time (in ms). The y-axis represents individual electrode positions. Starting from the bottom ofthe graph, the electrodes are divided into sections from posterior to anterior scalp. Within a section, the electrodes are arranged from theright most lateral to the left most lateral sites. The sections are labeled based on the midline of the scalp (e.g. "central" includeselectrodes over temporal scalp).
INSAR Russo et al./Auditory–somatosensory integration in ASD 11

years, will be informative with regard to the question of
early MSI in children.
The current data indicate that children with ASD do
not automatically combine sensory inputs early in the
processing hierarchy to the same degree as TD indivi-
duals, as assessed by the absence of audio-somatosensory
MSIs in the 100- to 200-ms time frame. These findings are
in line with several theories of ASD, including the
temporal binding hypothesis [Brock, Brown, & Boucher,
2002] and the underconnectivity hypothesis [e.g. Just,
Cherkassky, Keller, Kana, & Minshew, 2007]. Both of
these theories begin from the premise that in ASD there is
a tendency to focus on the local rather than the global
aspects of information, a thesis that was originally
formulated by Frith and Happe [1994] and termed as
weak central coherence. According to the temporal
binding hypothesis, the local bias in ASD is related to a
failure to integrate information from different specialized
networks in the brain [Brock et al., 2002]. This failure to
integrate information has also received some support
from findings of local overconnectivity between prox-
imal areas as evidenced by findings of increased mini-
columnar density in postmortem studies [Casanova,
2007] and functional underconnectivity between more
distal areas of the brain as evidenced by fMRI [Just,
Cherkassy, Keller, & Minshew, 2004]. Findings of both
brain under- and over- connectivity [Mizuno, Villalobos,
Davies, Dahl, & Muller, 2006] provide a simple physio-
logical basis for proposed differences in multisensory
processing in ASD, which would be expected to require
distal brain connectivity [e.g. Falchier, Clavagnier,
Barone, & Kennedy, 2002; Rockland & Ojima, 2003;
Smiley & Falchier, 2009]. Evidence of functional under-
connectivity between frontal and visual areas in ASD has
been replicated during both rest [Cherkassky, Kana,
Keller, & Just, 2006] and a variety of experimental tasks
that include response inhibition [Kana, Keller, Minshew,
& Just, 2007], working memory [Koshino et al., 2005],
and executive function [Just et al., 2007]. In addition to
findings of underconnectivity, functional overconnectivity
between the thalamus, a key relay area in sensory
processing, and certain frontal regions that include the
left insula and right postcentral and middle frontal gyri
has also been noted [Mizuno et al., 2006]. Pathophysio-
logical findings from postmortem tissue of the brains of
persons with ASD are also consistent with atypical
connectivity. There are reliable differences in relation to
the TD brain, with findings of a smaller minicolumnar
width, a greater number of minicolumns, and increased
neuronal density in various cortical areas including the
frontal and temporal lobes [Casanova, 2004, 2006].
Together these findings indicate possible functional
and anatomical differences that may contribute to
problems with MSI, which requires rapid and accurate
communication between subcortical and cortical areas.
Crucially, evidence from anatomical studies have shown
that there are direct long range connections between the
various sensory cortical areas [Falchier et al., 2002;
Rockland & Ojima, 2003] and it may be the disruption
of this form of connectivity that leads to impairment of
early MSI in ASD.
A major question raised is how to reconcile our
findings with the few behavioral studies on multisensory
processing in autism that suggest that the integration of
basic stimuli is typical in ASD [e.g. Bebko et al., 2006; van
der Smagt et al., 2007], whereas it is the integration of
more complex stimuli, such as language that is impaired
[e.g. Mongillo et al., 2008; Smith & Bennetto, 2007]. All
these studies have involved auditory–visual stimuli, so
one possibility is that the difference lies in the sensory
modalities involved. Another possibility is that while
individuals with autism are able to benefit from multi-
sensory inputs, especially for simpler stimulus and task
configurations, the underlying brain processes never-
theless differ.
It is also possible that for persons with ASD, actively
attending the stimuli is necessary for integration to
occur. That is, that MSI processes that occur in the
absence of directed attention in TD individuals require
attention to be achieved in this population. It will be
interesting to assess if making the stimuli task-relevant
serves to ‘‘normalize’’ the neural responses, or if typical
behavioral facilitation is seen in the face of different
neural processing.
Another potential contribution to differences in the
integration of the auditory and somatosensory stimuli
might come from the small but reliable differences in
the magnitude of the unisensory responses that were
observed between the groups. Although for both groups
there were robust unisensory responses which were in the
main highly similar between the groups, these appeared
slightly smaller in the ASD group. Just how this would
impact subsequent processing is difficult to say, but we
doubt that the whole-scale differences that we see in
multisensory processing would be mitigated by ‘‘normal-
izing’’ the unisensory response. One possibility is that in
ASD multisensory processing is more (or less) mature
than for TD individuals. Only by collecting considerably
more data points, however, will we be able to paint the
developmental trajectory of auditory–somatosensory MSI
and address such questions. A number of studies have
specifically examined how the ‘‘effectiveness’’ of uni-
sensory stimulation impacts MSI. For studies that record
single cell activity, effectiveness is assessed by the number
of spikes per unit of time. Such studies have demon-
strated that the more ineffective unisensory stimuli are at
eliciting a neuronal response when presented alone, the
greater the multisensory enhancement tends to be when
they are presented together [e.g. Stein & Stanford, 2008;
Stein, Stanford, Ramachandran, Perraullt, & Rowland,
12 Russo et al./Auditory–somatosensory integration in ASD INSAR

2009]. This is referred to as the ‘‘principle of inverse
effectiveness’’. According to this principle, one could in
fact make the case that the unisensory stimuli used here
were less effective for the ASD group and therefore that
their MSIs should in fact be stronger. The relationship
between the robustness of the unisensory response and
the magnitude of the multisensory response has not been
so straightforward when tested using other methods. For
example, some fMRI studies find support for inverse
effectiveness [e.g. Werner & Noppeney, 2009], whereas
others have actually shown quite the opposite, with
enhanced integration seen as the effectiveness of uni-
sensory stimulation increased [Kim & James, 2010].
Behavioral data from our laboratory reveal that for more
complex linguistic stimuli, the largest audiovisual multi-
sensory effects are present for stimuli that are at
intermediary stages of effectiveness, suggesting that there
is not necessarily a linear relationship between the
effectiveness of unisensory stimuli and MSI [e.g. Ma,
Zhou, Ross, Foxe, & Parra, 2009; Ross et al., 2007]. Clearly
much work remains to be done to understand how the
differences in unisensory processing relate to differences
in multisensory processing.
Conclusions
The findings from this study provide preliminary con-
firmatory evidence for anecdotal and clinical reports of a
difficulty in integrating information from multiple
sensory modalities among persons with ASD. Much work,
of course, remains to be done to fully understand not
only the differences in how multisensory inputs are
integrated but also how these differences affect higher-
order processing, and the impact that a lack of early
integration has on the pathogenesis of persons with
ASDs. In addition, it will be important to determine
whether there are specific phenotypic characteristics of
autism (e.g. sensory overresponsiveness) that are more
strongly associated with differences in multisensory
processing than others, as well as whether MSI deficits
are more strongly associated with specific subtypes of the
autism spectrum than others (e.g. autism vs. Asperger’s
syndrome).
Acknowledgments
Initial support for this work came from a pilot grant from
Cure Autism Now (J.J.F.). Additional support came from
the US National Institute of Mental Health (MH 085322
to S.M. and J.J.F.) and The Wallace Research Foundation
(S.M. and J.J.F.). NR received additional support from a
Post Doctoral Research Grant from the Fondation du
Quebec de Recherche sur la Societe et la Culture and from
the Autism Research Training Program through the
Canadian Institute of Health Research.
References
American Psychiatric Association. (2000). Diagnostic and statis-
tical manual of mental disorders: DSM-IV-TR. Washington,
DC: American Psychiatric Association.
Ayers, J.A. (1994). Sensory integration and the child. Los
Angeles, CA: Western Psychological Services.
Bebko, J.M., Weiss, J.A., Demark, J.L., & Gomez, P. (2006).
Discrimination of temporal synchrony in intermodal events
by children with autism and children with developmental
disabilities without autism. Journal of Child Psychology and
Psychiatry and Allied Disciplines, 47, 88–98.
Berman, A.L. (1961). Overlap of somatic and auditory cortical
response fields in anterior ectosylvian gyrus of cat. Journal of
Neurophysiology, 24, 595–607.
Bertone, A., Mottron, L., Jelenic, P., & Faubert, J. (2003). Motion
perception in autism: A ‘‘complex’’ issue. Journal of Cogni-
tive Neurosciences, 15, 218–225.
Bertone, A., Mottron, L., Jelenic, P., &, Faubert, J. (2005).
Enhanced and diminished visuo-spatial information proces-
sing in autism depends on stimulus complexity. Brain, 128,
2430–2441.
Brett-Green, B.A., Miller, L.J., Gavin, W.J., & Davies, P.L. (2008).
Multisensory integration in children: A preliminary ERP
study. Brain Research, 1242, 283–290.
Bonnel, A., Mottron, L., Peretz, I., Trudel, M., Gallun, E., &
Bonnel, A.M. (2003). Enhanced pitch sensitivity in indivi-
duals with autism: A signal detection analysis. Journal of
Cognitive Neuroscience, 15, 226–235.
Brock, J., Brown, C.C., & Boucher, J. (2002). The temporal
binding deficit hypothesis of autism. Development and
Psychopathology, 14, 209–224.
Carriere, B.N., Royal, D.W., Perrault, T.J., Morrison, S.P.,
Vaughan, J.W., et al. (2007). Visual deprivation alters the
development of cortical multisensory integration. Journal of
Neurophysiology, 98, 2858–2867.
Casanova, M.F. (2004). White matter volume increase and
minicolumns in autism. Annals of Neurology, 56, 453.
Casanova, M.F. (2006). Neuropathological and genetic findings
in autism: The significance of a putative minicolumnopathy.
Neuroscientist, 12, 435–441.
Casanova, M.F. (2007). The neuropathology of autism. Brain
Pathology, 17, 422–433.
Ceponiene, R., Rinne, T., & Naatanen, R. (2002). Maturation of
cortical sound processing as indexed by event-related poten-
tials. Clinical Neurophysiology, 113, 870–882.
Cesaroni, L., & Garber, M. (1991). Exploring the experience of
autism through firsthand accounts. Journal of Autism and
Developmental Disorders, 21, 303–313.
Cherkassky, V.L., Kana, R.K., Keller, T.A., & Just, M.A. (2006).
Functional connectivity in a baseline resting-state network in
autism. Neuroreport, 17, 1687–1690.
Crane, L., Goddard, L., & Pring, L. (2009). Sensory processing in
adults with autism spectrum disorders. Autism, 13, 215–228.
de Gelder, B., Vroomen, J., & Van der Heide, L. (1991). Face
recognition and lip-reading in autism. European Journal of
Cognitive Psychology, 3, 69–86.
Driver, J. (1996). Enhancement of selective listening by illusory
mislocation of speech sounds due to lip-reading. Nature, 381,
66–68.
INSAR Russo et al./Auditory–somatosensory integration in ASD 13

Falchier, A., Clavagnier, S., Barone, P., & Kennedy, H. (2002).
Anatomical evidence of multimodal integration in primate
striate cortex. Journal of Neuroscience, 22, 5749–5759.
Foxe, J.J., & Molholm, S. (2009). Ten years at the multisensory
forum: Musings on the evolution of a field. Brain Topography,
32, 149–154.
Foxe, J.J., & Schroeder, C.E. (2005). The case for feedforward
multisensory convergence during early cortical processing.
Neuroreport, 16, 419–423.
Foxe, J.J., Morocz, I.A., Murray, M.M., Higgins, B.A., Javitt, D.C.,
& Schoreder, C.E. (2000). Multisensory auditory-somatosen-
sory interactions in early cortical processing revealed by high-
density electrical mapping. Brain Research, Cognitive Brain
Research, 10, 77–83.
Foxe, J.J., Wyliw, G.R., Martinez, A., Schoeder, C.E., Javitt, D.C.,
et al. (2002). Autidory-somatosensory multisensory proces-
sing in auditory association cortex: An fMRI study. Journal of
Neurophyisology, 88, 540–543.
Frens, M.A., & Van Opstal, A.J. (1995). A quantitative study of
auditory-evoked saccadic eye movements in two dimensions.
Experimental Brain Resarch, 107, 103–117.
Frith, U., & Happe, F. (1994). Autism: Beyond ‘‘theory of mind’’.
Cognition, 50, 115–132.
Giard, M.H., & Peronnet, F. (1999). Auditory-visual interaction
during multimodal object recognition in humans: A beha-
vioral and electrophysiological study. Journal of Cognitive
Neurosciences, 11, 473–490.
Gori, M., Del Viva, M., Sandini, G., & Burr, D.C. (2008). Young
children do not integrate visual and haptic form information.
Current Biology, 18, 694–698.
Grandin, T. (1992). An inside view of autism. In: Schopler, E., &
Mesiboc, G.B., editors. High-functioning individuals with
autism. New York: Plenum Press, pp 105–126.
Guthrie, D., & Buchwald, J.S. (1991). Significance testing of
difference potentials. Psychophysiology, 28, 240–244.
Hermelin, B., & O’Connor, N. (1970). Psychological experiments
with autistic children. Oxford, England: Pergamon Press.
Hotting, K., Rosler, F., & Roder, B. (2004). Altered auditory-tactile
interactions in congenitally blind humans: An event-related
potential study. Experimental Brain Research, 159, 370–381.
Just, M.A., Cherkassy, V.L., Keller, T.A., & Minshew, N. (2004).
Cortical activation and synchronization during sentence
comprehension in high-functioning autism: Evidence of
underconnectivity. Brain, 127, 1811–1821.
Just, M., Cherkassky, V.L., Keller, T.A., Kana, R.K., & Minshew, N.J.
(2007). Functional and anatomical underconnectivity in
autism: Evidence from an fMRI study of an executive function
task and corpus collosum morphometry. Cerebral Cortex, 27,
951–961.
Kana, R.K., Keller, T.A., Minshew, N.J., & Just, M.A. (2007).
Inhibitory control in high-functioning autism: Decreased
activation and underconnectivity in inhibition networks.
Biological Psychiatry, 62, 198–206.
Kim, S., & James, T.W. (2010). Enhanced effectiveness in visuo-
haptic object-selective brain regions with increasing stimulus
salience. Human Brain Mapping, 31, 678–693.
Klin, A., Sparrow, S.S., de Bildt, A., Cichetti, D.V., Cohen, D.J., &
Volkmar, F.R. (1999). A normed study of face recognition in
autism and related disorders. Journal of Autism and Devel-
opmental Disorders, 29, 499–508.
Koshino, H., Carpenter, P.A., Minshew, N.J., Cherkassky, V.L.,
Keller, T.A., & Just, M.A. (2005). Functional connectivity in an
fMRI working memory task in high-functioning autism.
Neuroimage, 24, 810–821.
Lane, A.E., Young, R.L., Baker, A.E., & Angley, M.T. (2010).
Sensory processing subtypes in autism: Association with
adaptive behavior. Journal of Autism and Developmental
Disorders, 40, 112–122.
Lutkenhoner, B., Lammertmann, C., Simoes, C., & Hari, R.
(2002). Magnetoencephalographic correlates of audiotactile
interaction. Neuroimage, 15, 509–522.
Lewkowicz, D.J. (2002). Heterogeneity and heterochrony in the
development of intersensory perception. Brain Research:
Cognitive Brain Research, 14, 41–63.
Lord, C., Rutter, M., & Le Couteur, A. (1994). Autism diagnostic
interview-revised: A revised version of a diagnostic interview
for caregivers of individuals with possible pervasive develop-
mental disorders. Journal of Autism and Developmental
Disorders, 24, 659–685.
Lord, C., Rutter, M., DiLavore, P.C., & Risi, S. (1999). Autism
diagnostic observation schedule. Los Angeles, CA: Western
Psychological Services.
Lovaas, O.I., & Schreibman, L. (1971). Stimulus overselectivity of
autistic children in a two stimulus situation. Behaviour
Research and Therapy, 9, 305–310.
Lovaas, O.I., Schreibman, L., Koegel, R., & Rehm, R. (1971).
Selective responding by autistic children to multiple sensory
input. Journal of Abnormal Psychology, 77, 211–222.
Ma, W.H., Zhou, X., Ross, L.A., Foxe, J.J., & Parra, L.C. (2009). Lip-
reading aids word recognition most in moderate noise:
A Bayesian explanation using high-dimensional feature space.
PLoS One 4.e4638. DOI: 10.1371/journal.pone.0004638.
Magnee, M.J., de Gelder, B., van Engeland, H., & Kemner, C.
(2008). Audiovisual speech integration in pervasive develop-
mental disorder: Evidence from event related potentials.
Journal of Child Psychology and Psychiatry, 49, 995–1000.
Magnee, M.J., Oranje, B., van Engeland, H., Kahn, R.S., &
Kemner, C. (2009) Cross-sensory gating in schizophrenia and
autism spectrum disorder: EEF evidence for impaired brain
connectivity? Neuropsychologia, 7, 1728–1732.
Mizuno, A., Villalobos, M.E., Davies, M.M., Dahl, B.C., &
Muller, R.A. (2006) Partially enhanced thalamocortical func-
tional connectivity in autism. Brain Research, 1104, 160–174.
Molholm, S., Ritter, W., Murray, M.M., Javitt, D.C.,
Schroeder, C.E., & Foxe, J.J. (2002). Multisensory auditory-
visual interactions during early sensory processing in
humans: A high-density electrical mapping study. Brain
Research: Cognitive Brain Research, 14, 115–128.
Molholm, S., Ritter, W., Javitt, D.C., & Foxe, J.J. (2004). Multi-
sensory visual-auditory object recognition in humans: A high-
density electrical mapping study. Cerebral Cortex, 14, 452–465.
Molholm, S., Martinez, A., Shpaner, M., & Foxe, J.J. (2007).
Object-based attention is multisensory: Co-activation of an
object’s representations in ignored sensory modalities.
European Journal of Neurosciences, 26, 499–509.
Mongillo, E.A., Irwin, J.R., Whalen, D.H., Klaiman, C., Carter, A.S.,
& Schultz, R.T. (2008). Audiovisual processing in children with
14 Russo et al./Auditory–somatosensory integration in ASD INSAR

and without autism spectrum disorders. Journal of Autism and
Developmental Disorders, 38, 1349–1358.
Murray, M.M., Molholm, S., Michel, C.M., Heslenfeld, D.J.,
Ritter, W., et al. (2005). Grabbing your ear: Rapid auditory-
somatosensory multisensory interactions in low-level sensory
cortices are not constrained by stimulus alignment. Cerebral
Cortex, 15, 963–974.
Nardini, M., Jones, P., Bedford, R., & Braddick, O. (2008).
Development of cue integration in human navigation.
Current Biology, 18, 689–693.
O’Neill, M., & Jones, R.S.P. (1997). Sensory-perceptual abnorm-
alities in autism: A case for more research? Journal of Autism
and Developmental Disorders, 27, 283–293.
O’Riordan, M. (2004). Superior visual search in autism. Autism,
8, 229–248.
O’Riordan, M., & Plaisted, K. (2001). Enhanced discrimination in
autism. Quarterly Journal of Experimental Psychology, 54, 961–979.
O’Riordan, M., & Passetti, F. (2006). Discrimination in autism
within different modalities. Journal of Autism and Develop-
mental Disorders, 36, 665–675.
Ozonoff, S., & Strayer, D.L. (1997). Inhibitory function in non-
retarded children with autism. Journal of Autism and
Developmental Disorders, 27, 59–77.
Perrott, D.R., Saberi, K., Brown, K., & Strybel, T.Z. (1990).
Auditory psychomotor coordination and visual search per-
formance. Perceptual Psychophysics, 48, 214–226.
Putzar, L., Goerendt, I., Lange, K., Rosler, F., & Roder, B. (2007).
Early visual deprivation impairs multisensory interactions in
humans. Nature Neuroscience, 10, 1243–1245.
Rockland, K.S., & Ojima, H. (2003). Multisensory convergence in
calcarine visual areas in macaque monkey. International
Journal of Psychophysiology, 50, 19–26.
Ross, L.A., Saint-Amour, D., Leavitt, V.M., Molholm, S., Javitt, D.C.,
& Foxe, J.J. (2007). Impaired multisensory processing in
schizophrenia: Deficits in the visual enhancement of speech
comprehension under noisy environmental conditions. Schi-
zophrenia Research, 97, 173–183.
Schoen, S.A., Miller, L.J., Brett-Green, B.A., & Nielsen, D.M.
(2009). Physiological and behavioral differences in sensory
processing: A comparison of children with autism spectrum
disorder and sensory modulation disorder. Frontiers in
Integrative Neurosciences, 29 [Electronic version].
Schroeder, C.E., & Foxe, J.J. (2005). Multisensory contributions
to low-level ‘‘unisensory’’ processing. Current Opinions in
Neurobiology, 15, 454–458.
Shams, L., Kamitani, Y., & Shimojo, S. (2000). Illusions: What
you see is what you hear. Nature, 14, 788.
Shams, L., Kamitani, Y., & Shimojo, S. (2002). Visual illusion
induced by sound. Brain Research -Cognitive Brain Research,
14, 147–152.
Smith, E.G., & Bennetto, L. (2007). Audiovisual speech integra-
tion and lipreading in autism. Journal of Child Psychiatry
and Psychology, 48, 813–821.
Smiley, J.F., & Falchier, A. (2009). Multisensory connections of
monkey auditory cerebral cortex. Hearing Research, 258,
37–46.
Spence, C., & Driver, J. (1997). On measuring selective attention
to an expected sensory modality. Perceptual Psychophysics,
59, 389–403.
Sperdin, H.F., Cappe, C., Foxe, J.J., & Murray, M.M. (2009). Early
low-level auditory-somatosensory multisensory interaction
impact reaction time speed. Frontiers in Integrative Neuros-
ciences, 3 [Epub March 11].
Stein, B.E., & Meredith, M.A. (1990). Multisensory integration.
Neural and behavioral solutions for dealing with stimuli from
different sensory modalities. Annuals of the New York
Academy of Science, 608, 51–65; discussion 65–70.
Stein, B.E., & Stanford, T.R. (2008). Multisesnory integration:
Current issues from the perspective of the single neuron.
Nature Neuroscience Reviews, 9, 255–266.
Stein, B.E., Stanford, T.R., Ramachandran, R., Perrault, Jr., T.J., &
Rowland, B.A. (2009). Challenges in quantifying multisen-
sory integration: Alternative criteria, models and inverse
effectiveness. Experimental Brain Research, 198, 113–126.
Stroh, G., & Buick, D. (1964). Perceptual development and childhood
psychosis. British Journal of Medical Psychology, 37, 291–299.
Tager-Flusberg, H. (1996). Brief report: Current theory and
research on language and communication in autism. Journal
of Autism and Developmental Disorders, 26, 169–172.
Tager-Flusberg, H., & Sullivan, K. (1994). A second look at
second-order belief attribution in autism. Journal of Autism
Developmental Disorder, 24, 577–586.
Talsma, D., & Woldorff, M.G. (2005). Selective attention and
multisensory integration: Multiple phases of effects on the evoked
brain activity. Journal of Cognitive Neuroscience, 17, 1098–1114.
Taylor-Clarke, M., Kennett, S., & Haggard, P. (2002). Vision
modulates somatosensory cortical processing. Current Biology,
12, 233–236.
Teder-Salejarvi, W., McDonald, J.J., Di Russo, F., & Hillyard, S.A.
(2002). An analysis of audio-visual crossmodal integration by
means of event-related potential (ERP) recordings. Brain
Research Cognitive Brain Research, 14, 106–114.
Tomchek, S.D. (2007). Sensory processing in children with and
without autism: A comparative study using the short sensory
profile. American Journal of Occupational Therapy, 61,
190–200.
van der Smagt, M.J., van Engeland, H., & Kemner, C. (2007). Can
you see what is not there? Low-level auditory-visual integra-
tion in autism spectrum disorder. Journal of Autism and
Developmental Disorders, 37, 2014–2019.
Vroomen, J., & Stekelenburg, J.J. (2010). Visual anticipatory
information modulates multisensory interaction of artificial
audiovisual stimuli. Journal of Cognitive Neurosciences, 22,
1583–1596.
Wallace, M.T., Perrault, Jr., T.J., Hairston, W.D., & Stein, B.E.
(2004). Visual experience is necessary for the development of
multisensory integration. Journal of Neurosciences, 24,
9580–9584.
Wechsler, D. (1999). Wechsler abbreviated scale of intelligence.
San Antonio, TX: Psychological Corporation.
Werner, S., & Noppeney, U. (2010). Superadditive responses in
superior temporal sulcus predict audiovisual benefits in
object categorization. Cerebral Cortex, 20, 1829–1842.
Williams, D. (1994). Somebody somewhere. London: Doubleday.
Zahn, J.R., Abel, L.A., & Dell’Osso, L.F. (1978). Audio-ocular
response characteristics. Sensory Processes, 2, 32–37.
INSAR Russo et al./Auditory–somatosensory integration in ASD 15





























