Embed Size (px)
Citation preview

Neurosciences
Réorganisation du cortex sensorimoteur dans le cadre de la paralysiecérébrale unilatérale : apports des neurosciences
Reorganization of the sensorimotor cortex in children with spastic
unilateral cerebral palsy: What can we learn from neuroscience?
M. Dinomais a,b,*, L. Hertz-Pannier c, S. Nguyen The Tich a,d
a EA4094, laboratoire d’ingénierie des systèmes automatisés (LISA), LUNAM, université d’Angers, 62, avenue Notre-Dame-du-Lac,49000 Angers, France
b Département de médecine physique et de réadaptation, centre les Capucins, CHU–CRRRF, 28, rue des Capucins, BP 40329,49103 Angers cedex 02, France
c U663, Inserm-CEA, neurospin-UNIACT, CEA-SACLAY, route Nationale, 91400 Gif-sur-Yvette, Franced Département de neuropédiatrie, LUNAM, université d’Angers, CHU d’Angers, 4, rue Larrey, 49000 Angers, France
Disponible sur Internet le 6 fevrier 2014
Résumé
La paralysie cérébrale (PC) est définie comme un trouble permanent du mouvement secondaire à une lésion cérébrale nonprogressive survenue pendant la période fœtale ou lors des deux premières années de vie. Lorsque la lésion concerne uniquement unhémisphère, on parle de PC unilatérale. Dans le cadre de la PC unilatérale, les études utilisant la stimulation magnétiquetranscrânienne et l’IRM fonctionnelle ont montré que la plasticité post-lésionnelle du contrôle centrale de la motricité (motricité-cérébrale) pouvait se réaliser selon deux schémas. Certains patients, présentant des lésions majeures du faisceau corticospinal,développent un faisceau pyramidal ipsilatéral à l’hémisphère sain. Via ce faisceau ipsilatéral, le cortex moteur de l’hémisphère saincontrôle la main parétique (main ipsilatérale à l’hémisphère sain). Chez ces sujets, on objective le cortex moteur primaire au niveaudu gyrus précentral de l’hémisphère sain. Ce type de réorganisation ipsilatérale est le plus souvent observé dans les lésionspériventriculaires avec des dommages du faisceau pyramidal dans la substance blanche, et se rencontre uniquement dans le cadre dela PC. À l’opposé, chez des patients présentant un accident vasculaire cérébral néonatal de l’artère cérébrale moyenne, le cortexmoteur primaire reste situé au niveau du gyrus précentral de l’hémisphère lésé, c’est-à-dire « controlatéralement » à la mainparétique. Indépendamment de cette réorganisation intra- ou inter-hémisphérique du cortex moteur, le cortex sensitif primaire estquant à lui toujours retrouvé au niveau de l’hémisphère lésé (gyrus post-central). Ces deux types de réorganisation de la motricité-cérébrale peuvent influencer l’efficacité clinique des thérapies utilisées en rééducation.# 2013 Elsevier Masson SAS. Tous droits réservés.
Abstract
Cerebral palsy (CP) is a non-progressive injury to the developing central nervous system and defines as permanent disorders ofthe development of movement and posture, causing activity limitation. This neurodevelopmental disorder may lead to spasticunilateral cerebral palsy after early unilateral brain lesions. Physical and rehabilitation medicine has a particular interest in the study
www.em-consulte.com
Disponible en ligne sur www.sciencedirect.com
ScienceDirect
Motricité cérébrale 35 (2014) 3–14
* Auteur correspondant.Adresse e-mail : [email protected] (M. Dinomais).
0245-5919/$ – see front matter # 2013 Elsevier Masson SAS. Tous droits réservés.
http://dx.doi.org/10.1016/j.motcer.2013.12.001
of organization and reorganization of the sensorimotor cortex following early brain injury. From neuroscience standpoint, earlybrain lesions have been shown to induce substantial neural reorganization owing to the higher plasticity in the developing brain.Unilateral injuries either to the motor cortex or the corticospinal tract can lead to different patterns of reorganization of thesensorimotor cortex. Many patients develop ipsilateral corticospinal pathways to control the paretic hand with the non-lesionedhemisphere. This type of reorganization is often observed following unilateral periventricular brain lesions, which damage thecorticospinal tracts in the periventricular white matter. In this group of patients, the primary motor cortex has been found to berepresented in the non-lesioned precentral gyrus ipsilateral to the paretic side. Inversely, in patients with perinatal unilateral middlecerebral artery stroke, primary motor cortex remains organised in the lesioned precentral gyrus contralateral to the paretic hand.However, regardless of these inter- or intrahemispheric motor representations, the primary somatosensory cortex representationremains in the lesioned hemisphere in both groups. These two types of corticospinal reorganization could influence the efficacy ofrehabilitation.# 2013 Elsevier Masson SAS. All rights reserved.
Mots clés : Paralysie cérébrale ; Cortex moteur ; Cortex somesthésique ; Plasticité cérébrale
Keywords: Cerebral palsy; Motor cortex; Sensory cortex; Plasticity
M. Dinomais et al. / Motricité cérébrale 35 (2014) 3–144
1. Introduction
Le professeur André Grossiord (père fondateur de laMédecine Physique et Réadaptation en France) sequestionnait en 1980 dans l’éditorial du premier numérode la revue Motricité Cérébrale [1] à propos de la« neuro-rééducation » dans les pathologies neurologi-ques d’origine centrale : « Au-delà de l’étiologie, de lacause première vasculaire, anoxique, mécanique, ne setrouve-t-on pas devant des faits physiopathologiquesvoisins ? ». Il existe donc un intérêt particulier de laMPR pour l’étude de l’organisation et de la réorganisa-tion cérébrale dans les suites d’une lésion del’encéphale, afin d’appréhender au mieux les méca-nismes neurophysiologiques de plasticité cérébrale et infine d’orienter nos prises en charge rééducatives.
La plasticité cérébrale est un « processus continupermettant des modifications à moyen et long termes del’organisation synaptique pour une meilleure efficacitédes réseaux neuronaux » [2]. On décrit une plasticité« naturelle » qui comprend « aussi bien le développe-ment cérébral normal et le maintien des circuitsneuronaux adultes » [2] et une plasticité « post-lésionnelle » qui est la réorganisation neuronale etsynaptique faisant suite à une lésion de l’encéphale. Lescapacités de plasticité post-lésionnelle du cerveau sontdéterminées par des mécanismes moléculaires intra- etextracellulaires. Ces mécanismes sont régulés par desprocessus dépendants de l’activité cellulaire « activitydependent processes » et par les interactions avec lemilieu extérieur (« interaction with the external world »[3]). Des interventions externes (pharmacologiques et/ou physiques), en stimulant l’activité cellulaire auniveau des zones cérébrales lésées et périlésionnelles,semblent pouvoir favoriser un certain degré de plasticité
post-lésionnelle dans des proportions non connues pourle moment [3].
La connaissance des mécanismes de plasticités(naturelle et post-lésionnelle) dans le système senso-rimoteur est nécessaire lorsque l’on veut développer desmodèles de prise en charge des déficiences sensorimo-trices. Du fait de la complexité du développementcérébral et de la maturation, il apparaît que laréorganisation du système sensorimoteur après lésioncérébrale ne peut pas être conçue comme identiqueentre des lésions intervenant tôt dans la vie de l’individu(fœtus, enfance) et des lésions survenant sur un cerveau« adulte ». Il a été montré que les incapacités motricesissues de lésions cérébrales apparaissent variables enfonction du moment de survenue de ces lésions. Eneffet, chez le singe [4], à lésion de taille égale et delocalisation identique (au niveau du gyrus précentral), larécupération motrice est meilleure pour des lésionsintervenues tôt dans la vie (en période néonatale)comparées à des lésions plus tardives. De plus, plus lalésion est tardive (dans l’enfance) et plus les déficitss’apparentent à des déficits observés dans le cadre delésions survenues chez le singe adulte. Il apparaît alorsque les conséquences en termes de déficiences sontdifférentes selon le « timing » (moment de survenue dela lésion). La survenue d’une lésion sur un cerveauencore immature (que l’on appelle lésion cérébraleprécoce) semble de meilleur pronostic fonctionnelmoteur qu’une lésion plus tardive. Le rôle de l’âge dansla plasticité, au moins concernant le système moteur, estappelé « principe de Kennard » [4].
La paralysie cérébrale (PC) est définie [5] comme untrouble moteur permanent, secondaire à une lésioncérébrale, non évolutive, survenue, avant l’âge de 2 ans(seuil arbitraire), sur un cerveau en développement

M. Dinomais et al. / Motricité cérébrale 35 (2014) 3–14 5
(cerveau immature). On ne peut établir formellement lediagnostic de PC qu’à partir de l’âge de 4 ans. Le termePC regroupe donc tous les enfants et adultes souffrantd’une incapacité motrice, dans les suites d’une lésioncérébrale précoce non-évolutive, entraînant une res-triction de participation. Nous donnons ici la définitionanglaise tirée de Rosenbaum en 2007 [6] : (cerebralpalsy has been defined as) « a group of permanentdisorders of the development of movement and posture,causing activity limitation, that are attributed to non-progressive disturbances that occurred in the develop-ing fetal or infant brain ». Cette définition de la PCregroupe les termes francophones d’infirmité motricecérébrale (IMC) et « infirmité motrice d’originecérébrale » (IMOC). La PC est actuellement la causela plus commune de handicap moteur de l’enfant.L’incidence de cette pathologie est de 2 à 3/1000 naissances [7].
Les formes cliniques de PC sont décrites en fonctionde leur symptomatologie neurologique prédominante.Ainsi, on décrit des formes spastiques uni ou bilatérales.Ces formes représentent 85 % des formes de PC. Lesautres formes possibles sont : les formes dyskinétiquesqui regroupent les formes choréiques et dystoniques (5 à10 %), les formes ataxiques (5 %) qui sont souventaccompagnées d’hypotonie, et enfin les formes mixtesqui peuvent associer plusieurs composantes neurologi-ques (dystonies, spasticité. . .).
Les formes spastiques de PC peuvent être sub-divisées en fonction de la distribution des localisationstopographiques de l’atteinte motrice. On décrit :
� l’hémiplégie (hemiplegia) : l’atteinte motrice toucheun hémicorps, parfois aussi la face et prédominesouvent au membre supérieur. La marche est souventacquise vers 2 ans et permet à l’enfant une autonomiemotrice. Parfois, une atteinte controlatérale minimeexiste car le mécanisme responsable de l’atteinte aprovoqué une lésion qui n’est pas strictement limitéeà un hémisphère cérébral. Cette forme est appeléediplégie asymétrique (asymmetrical diplegia). C’estune forme frontière entre l’hémiplégie et la diplégie ;� la diplégie spastique (diplegia, ou anciennement
maladie de Little) : l’atteinte est diffuse maisprédomine sur les membres inférieurs avec une attitudemotrice caractéristique : rotation interne des hanches,flexion des genoux, pieds équins. Les membressupérieurs sont relativement épargnés, même s’ilexiste fréquemment des troubles du graphisme ;� la quadriplégie (quadriplegia) : l’atteinte est massive,
elle touche les quatre membres y compris le tronc. Ilexiste une insuffisance posturale de tronc et un
contrôle de la tête souvent faible qui empêchel’acquisition de la marche et de la station assiseautonome. Les membres supérieurs sont autantatteints que les membres inférieurs ;� la triplégie (triplegia) : trois membres sont atteints,
classiquement les deux membres inférieurs et unmembre supérieur ;� la monoplégie (monoplégia) : un seul membre est
atteint (membre supérieur ou inférieur). Cette formeest rare (1 % des cas de PC).
La PC spastique unilatérale constitue un modèled’étude de la réorganisation cérébrale et des moyens decompensation des déficiences après lésion sur uncerveau en développement [8]. Le mécanisme quisous-tend la plasticité cérébrale post-lésionnelle, per-mettant le transfert de fonctions du cortex cérébral lésévers des régions cérébrales intactes, a été beaucoupétudié chez l’adulte présentant un accident vasculairecérébral (AVC) mais encore peu chez l’enfantprésentant une PC.
La suite de cet article concerne essentiellement ladescription non exhaustive (il ne s’agit pas d’une revuede la littérature) des connaissances actuelles sur laréorganisation de la motricité-cérébrale après lésioncérébrale précoce dans le cadre de la paralysie cérébraleunilatérale. L’expression « motricité-cérébrale » ren-voie à l’ensemble des structures et réseaux cérébrauxqui interviennent dans la réalisation du mouvement(acte moteur).
2. Plasticité naturelle de la motricité-cérébrale :mise en place du faisceau corticospinal croisé
Avant d’étudier spécifiquement la plasticité post-lésionnelle de la motricité-cérébrale, il semble impor-tant d’exposer ici la mise en place du faisceaucorticospinal croisé. Durant la vie embryonnaire, lesprojections corticospinales naissent à partir du cortexmoteur et se développent de façon centrifuge (du centrevers la périphérie) pour gagner les motoneurones de lamoelle épinière (Fig. 1). À partir d’un hémisphèrecérébral (à partir du cortex moteur) naissent deuxprojections corticospinales pour chaque hémi-moelle. Àpartir de la vingtième semaine de gestation, lesfaisceaux cortico-spinaux descendants ont atteint leurcible [9] et débutent le processus de synaptogenèse(création de synapses) avec les motoneurones a de lacorne antérieure à chaque niveau médullaire segmen-taire. Durant cette phase, il est important de noter quechaque hémisphère cérébral envoie des projectionsbilatérales. Il existe donc initialement un faisceau
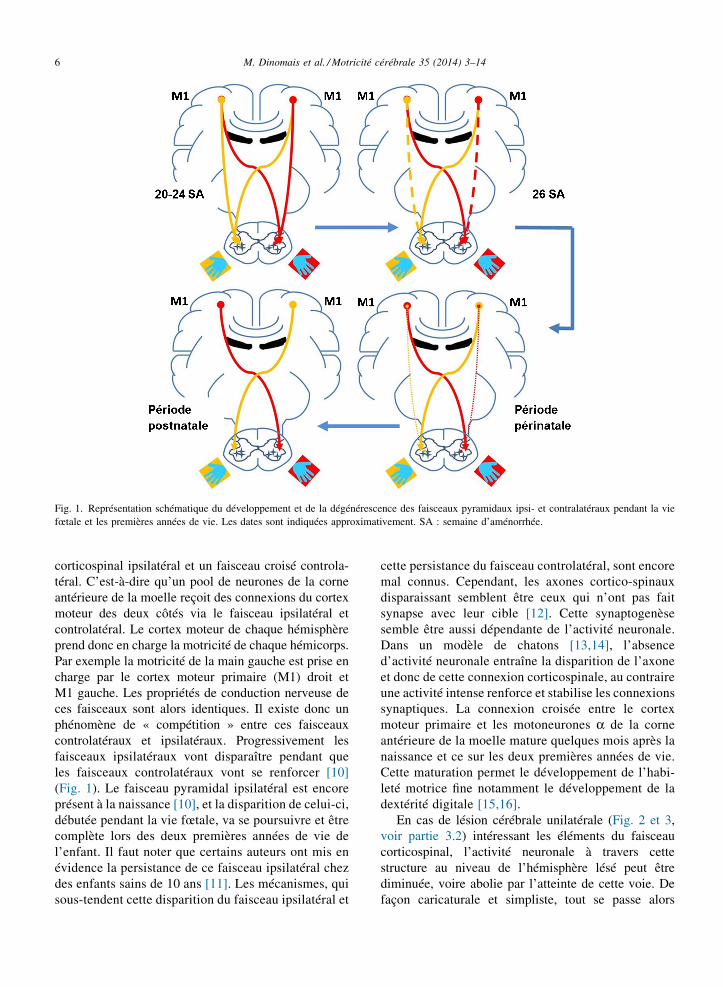
M. Dinomais et al. / Motricité cérébrale 35 (2014) 3–146
Fig. 1. Représentation schématique du développement et de la dégénérescence des faisceaux pyramidaux ipsi- et contralatéraux pendant la viefœtale et les premières années de vie. Les dates sont indiquées approximativement. SA : semaine d’aménorrhée.
corticospinal ipsilatéral et un faisceau croisé controla-téral. C’est-à-dire qu’un pool de neurones de la corneantérieure de la moelle reçoit des connexions du cortexmoteur des deux côtés via le faisceau ipsilatéral etcontrolatéral. Le cortex moteur de chaque hémisphèreprend donc en charge la motricité de chaque hémicorps.Par exemple la motricité de la main gauche est prise encharge par le cortex moteur primaire (M1) droit etM1 gauche. Les propriétés de conduction nerveuse deces faisceaux sont alors identiques. Il existe donc unphénomène de « compétition » entre ces faisceauxcontrolatéraux et ipsilatéraux. Progressivement lesfaisceaux ipsilatéraux vont disparaître pendant queles faisceaux controlatéraux vont se renforcer [10](Fig. 1). Le faisceau pyramidal ipsilatéral est encoreprésent à la naissance [10], et la disparition de celui-ci,débutée pendant la vie fœtale, va se poursuivre et êtrecomplète lors des deux premières années de vie del’enfant. Il faut noter que certains auteurs ont mis enévidence la persistance de ce faisceau ipsilatéral chezdes enfants sains de 10 ans [11]. Les mécanismes, quisous-tendent cette disparition du faisceau ipsilatéral et
cette persistance du faisceau controlatéral, sont encoremal connus. Cependant, les axones cortico-spinauxdisparaissant semblent être ceux qui n’ont pas faitsynapse avec leur cible [12]. Cette synaptogenèsesemble être aussi dépendante de l’activité neuronale.Dans un modèle de chatons [13,14], l’absenced’activité neuronale entraîne la disparition de l’axoneet donc de cette connexion corticospinale, au contraireune activité intense renforce et stabilise les connexionssynaptiques. La connexion croisée entre le cortexmoteur primaire et les motoneurones a de la corneantérieure de la moelle mature quelques mois après lanaissance et ce sur les deux premières années de vie.Cette maturation permet le développement de l’habi-leté motrice fine notamment le développement de ladextérité digitale [15,16].
En cas de lésion cérébrale unilatérale (Fig. 2 et 3,voir partie 3.2) intéressant les éléments du faisceaucorticospinal, l’activité neuronale à travers cettestructure au niveau de l’hémisphère lésé peut êtrediminuée, voire abolie par l’atteinte de cette voie. Defaçon caricaturale et simpliste, tout se passe alors
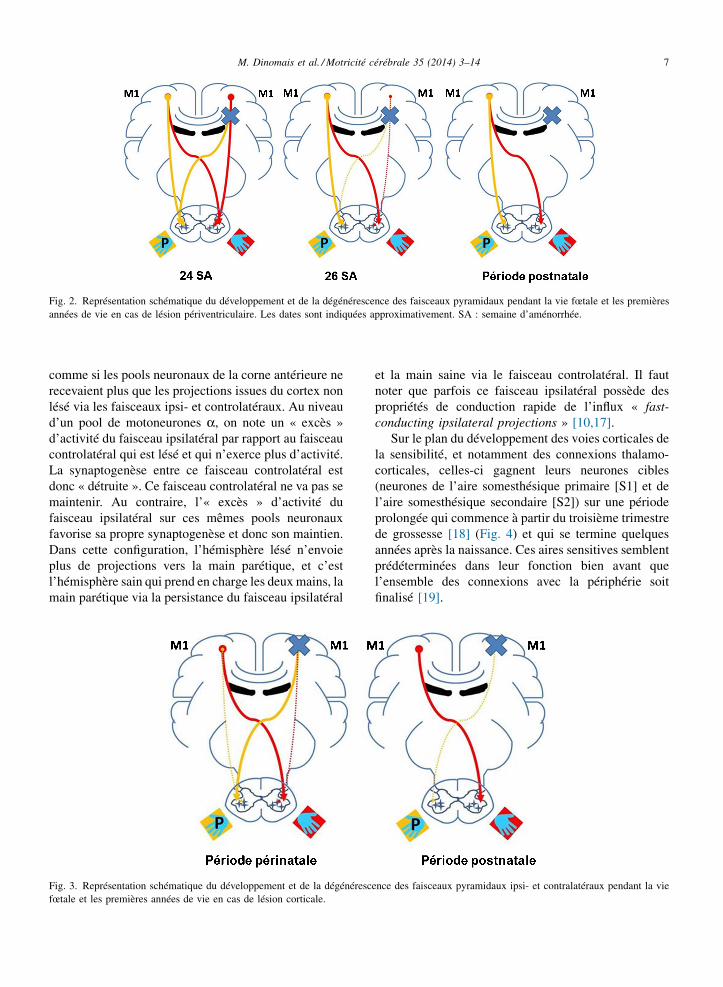
M. Dinomais et al. / Motricité cérébrale 35 (2014) 3–14 7
Fig. 2. Représentation schématique du développement et de la dégénérescence des faisceaux pyramidaux pendant la vie fœtale et les premièresannées de vie en cas de lésion périventriculaire. Les dates sont indiquées approximativement. SA : semaine d’aménorrhée.
comme si les pools neuronaux de la corne antérieure nerecevaient plus que les projections issues du cortex nonlésé via les faisceaux ipsi- et controlatéraux. Au niveaud’un pool de motoneurones a, on note un « excès »d’activité du faisceau ipsilatéral par rapport au faisceaucontrolatéral qui est lésé et qui n’exerce plus d’activité.La synaptogenèse entre ce faisceau controlatéral estdonc « détruite ». Ce faisceau controlatéral ne va pas semaintenir. Au contraire, l’« excès » d’activité dufaisceau ipsilatéral sur ces mêmes pools neuronauxfavorise sa propre synaptogenèse et donc son maintien.Dans cette configuration, l’hémisphère lésé n’envoieplus de projections vers la main parétique, et c’estl’hémisphère sain qui prend en charge les deux mains, lamain parétique via la persistance du faisceau ipsilatéral
Fig. 3. Représentation schématique du développement et de la dégénérescfœtale et les premières années de vie en cas de lésion corticale.
et la main saine via le faisceau controlatéral. Il fautnoter que parfois ce faisceau ipsilatéral possède despropriétés de conduction rapide de l’influx « fast-conducting ipsilateral projections » [10,17].
Sur le plan du développement des voies corticales dela sensibilité, et notamment des connexions thalamo-corticales, celles-ci gagnent leurs neurones cibles(neurones de l’aire somesthésique primaire [S1] et del’aire somesthésique secondaire [S2]) sur une périodeprolongée qui commence à partir du troisième trimestrede grossesse [18] (Fig. 4) et qui se termine quelquesannées après la naissance. Ces aires sensitives semblentprédéterminées dans leur fonction bien avant quel’ensemble des connexions avec la périphérie soitfinalisé [19].
ence des faisceaux pyramidaux ipsi- et contralatéraux pendant la vie

M. Dinomais et al. / Motricité cérébrale 35 (2014) 3–148
Fig. 4. Représentation schématique du développement des voies corticales de la somesthésie pendant la vie fœtale et les premières années de vie.
3. Réorganisation de la motricité-cérébraleaprès lésion cérébrale précoce
Bien qu’il soit clairement démontré qu’il n’existe pasde mécanismes de réparation neuronale possible, ilexiste des phénomènes de plasticité post-lésionnelleaprès lésion cérébrale précoce qui peuvent permettre deminimiser les déficiences consécutives à cet événementpathologique. Dans cette partie nous évoquerons laplasticité post-lésionnelle en général sans chercher àdiscuter du caractère inter- ou intra-modalité de cettedernière.
3.1. Réorganisation sur le plan anatomique
Sur le plan de l’anatomie structurelle, chez un mêmesujet, les lésions combinées de la substance blanche etde la substance grise entraînent des difficultés motricesau niveau de la main qui sont plus importantescomparées à des sujets souffrant de lésions cérébralesuniquement périventriculaires ou uniquement corticales[20]. De la même façon, chez des enfants ayant uneforme de PC unilatérale spastique, le degré d’atteintedes lésions périventriculaires semble prédire la fonctionmotrice. Ainsi plus la réduction de substance blancheest importante et plus la fonction motrice de la main estmauvaise [21]. Par ailleurs, après un AVC néonatal,l’atteinte du faisceau corticospinal est un facteurprédictif d’apparition d’une PC unilatérale (du côtéopposé à l’atteinte) [22].
Lee et al. [23] ont démontré, chez des enfants avecune PC bilatérale spastique liée à des lésions périven-triculaires, que l’intégrité et le respect des propriétés desfaisceaux pyramidaux (mesurée par imagerie dediffusion) étaient corrélés au degré de performance
motrice. Dans cette étude [23], il n’avait pas étéretrouvé de lien entre le volume de substance blanche etla fonction motrice.
En écho à ces recherches, Duque et al. [24] ontmontré que le degré d’asymétrie de taille des faisceauxcortico-spinaux mesurée au niveau des pédonculescérébraux était fortement corrélé avec la dextéritémanuelle chez des sujets atteints de PC. Plus il existeune asymétrie importante de ces pédoncules, en d’autrestermes plus un pédoncule est petit comparativement aupédoncule controlatéral, plus la dextérité est mauvaise,soulignant ainsi l’importance de l’intégrité de cefaisceau dans la motricité. Dans le cadre de lésionspériventriculaires bilatérales, le degré d’atteinte dutractus pyramidal est aussi corrélé avec la sévérité de lafonction motrice. Ces données sont à mettre en lien avecles données de tractographie et d’imagerie en diffusionqui ont montré que l’intégrité du faisceau pyramidalcontrolatéral (croisé) est un facteur essentiel pour lapréservation de la fonction motrice de la main parétiquechez l’enfant avec PC [25], ou après AVC périnatals[26].
L’ensemble de ces données montre l’importance dela préservation du faisceau corticospinal controlatéraldans la motricité-cérébrale, et souligne l’importance durespect de l’intégrité de celui-ci pour permettre unebonne fonction motrice.
3.2. Plasticité de la motricité-cérébrale et dufaisceau pyramidal
3.2.1. Réorganisation ipsilatéraleCe type d’adaptation que l’on appelle réorganisation
ipsilatérale peut se voir lors des lésions intervenues audébut du troisième trimestre de grossesse ou pendant la

M. Dinomais et al. / Motricité cérébrale 35 (2014) 3–14 9
période périnatale jusque dans le premiers mois de vie[27,28], voire jusque dans les deux premières années devie [29] (Fig. 2). Un tel type de réorganisation n’est pasdécrit lorsque la lésion cérébrale intervient après l’âgede deux ans [30]. Ce type de réorganisation est donccaractéristique de la PC, majoritairement dans le cadredes lésions périventriculaires.
En effet, sur le plan anatomique, le faisceaupyramidal traverse la substance blanche périventricu-laire à partir du gyrus précentral et post-central pourgagner la capsule interne. Ainsi, il est plus fréquentd’observer des atteintes très importantes allant jusqu’àl’interruption du faisceau corticospinal lors des lésionspériventriculaires de la substance blanche (lésions dudébut du troisième trimestre de grossesse, lésions de laprématurité), que lors de lésions corticales (qui ontmoins de probabilité d’atteindre le faisceau pyramidal).Même en cas de lésion importante cortico-sous-corticale, le faisceau pyramidal est souvent partielle-ment préservé [31]. Aussi la grande majorité des cas, oùon observe une adaptation ipsilatérale du faisceaupyramidal, est au cours des lésions périventriculaires dela substance blanche. Il est beaucoup plus rared’observer cette réorganisation dans le cas des lésionscorticales et/ou sous-corticales de la fin du troisièmetrimestre de grossesse et/ou de la période périnatale[32], et ce du fait, de la moindre probabilité d’atteinte dufaisceau pyramidal et de la disparition progressive desfaisceaux ipsilatéraux à cette période du développementcérébral.
Toujours dans le cadre des réorganisations ipsila-térales, certains patients montrent au niveau de lamotricité manuelle des pinces et des prises tout à faitfonctionnelles. Inversement, d’autres patients, ayant cemême type de réorganisation, possèdent une fonctionmotrice manuelle quasi-inexistante. Ce type de réorga-nisation ipsilatérale n’est jamais totalement efficace, etil n’existe pas de description de fonction manuellenormale en présence d’une telle réorganisation [30].
Une explication, probablement partielle, à cettevariabilité de la fonction motrice pour un même type deréorganisation et de lésion, est probablement que lespropriétés fonctionnelles (notamment les propriétés deconduction électro-physiologique de l’influx nerveux)du faisceau ipsilatéral sont dépendantes du moment oùest intervenue la lésion cérébrale. Il semble que dans lecadre d’une réorganisation ipsilatérale, plus tôt estintervenue, dans la vie fœtale, la lésion cérébrale,meilleur est le pronostic fonctionnel [32]. En effet, plustôt dans la vie fœtale, le faisceau ipsilatéral possèdeencore des propriétés satisfaisantes de conduction del’influx nerveux issu du cortex. Chez des enfants dont la
lésion cérébrale est intervenue autour de la naissance ouen post-natal, et qui possèdent une réorganisationipsilatérale de leur faisceau pyramidal, aucune motricitéfonctionnelle de la main parétique n’est observée[27,32] puisqu’il semble que le faisceau ipsilatéral nesoit plus assez fonctionnel à ce moment.
Sur le plan clinique, on note, chez les enfants ayantune réorganisation ipsilatérale, la présence de mouve-ments miroirs qui persistent après l’âge de 10 ans(période jusqu’à laquelle on peut voir des mouvementsmiroirs chez l’enfant sans critère pathologique) [11].Les mouvements miroirs sont des mouvements invo-lontaires de la main controlatérale à la main qui estintentionnellement bougée par le sujet. L’explication dece type de mouvement des deux mains alors qu’uneseule est volontairement bougée par le sujet semblerelativement intuitive dans le cadre d’une réorganisa-tion ipsilatérale après PC. En effet dans ce type deréorganisation motrice, le même cortex sain (non lésé)prend en charge les deux mains via le faisceaucontrolatéral et ipsilatéral. Or, l’inhibition motrice,notamment de M1 sur son homologue controlatéral, nepeut pas se produire. On peut imager cette explicationde la manière suivante. Le message moteur envoyé parle cortex moteur sain pour générer un mouvement del’une ou l’autre main emprunte les deux voiespyramidales en même temps sans qu’aucune inhibitionmotrice ne puisse s’exercer sur une des voies. Le messagemoteur arrive donc aux deux mains. Ceci a pourconséquences d’engendrer en même temps un mouve-ment dans les deux mains. On ne peut bien évidemmentpas exclure ici les autres hypothèses formulées dans lalittérature concernant la pathogénie des mouvementsmiroirs qui peut être liée à un défaut d’inhibition inter-hémisphérique entre les deux cortex moteurs ou à unealtération de la planification et de l’exécution motrice lorsde mouvements bi-manuels avec une activation bilatéralede M1 (pour une revue voir [33]).
En imagerie fonctionnelle, chez l’enfant avec PC etmanifestant une réorganisation ipsilatérale de leurfaisceau pyramidal, on note, lors de mouvement actifde la main parétique, des activations au niveau du cortexmoteur sain (comme lors du mouvement de la mainsaine) alors qu’il n’existe pas d’activation au niveau ducortex moteur lésé controlatéral au mouvement. Cetteactivation ipsilatérale au mouvement de M1 et cetteabsence d’activation controlatérale au niveau deM1 s’expliquent facilement par ce qui a été décrit ci-dessus. En effet, M1 de l’hémisphère sain prend encharge les deux mains alors que M1 lésé, en l’absencede faisceau pyramidal partant de cette zone, ne prendplus en charge la motricité de l’hémicorps controlatéral.

M. Dinomais et al. / Motricité cérébrale 35 (2014) 3–1410
Le mouvement de la main parétique entraîne donc uneactivation de M1 situé au niveau de l’hémisphère sain.
3.2.2. Réorganisation controlatéraleLa réorganisation controlatérale du faisceau pyra-
midal est une réorganisation comme on peut la voir chezl’adulte après un AVC, c’est-à-dire que la mainparétique est prise en charge par le cortex moteurlésé (cortex controlatéral) via le faisceau corticospinalcroisé (Fig. 3). Chaque hémisphère prend donc encharge la motricité de l’hémicorps controlatéral, et lesfaisceaux ipsilatéraux n’existent plus ou ne sont plusfonctionnels. Dans le cadre de la PC, ce typed’adaptation se voit essentiellement après une lésioncorticale et/ou sous-corticale, ce qui se rencontresurtout en toute fin de grossesse ou dans les premiersmois de vie après un AVC.
Sur le plan de l’analyse du cortex moteur, peud’études existent chez l’enfant après accident vasculairecérébral mais il semble que plus l’activité neuronale estpréservée au niveau de l’hémisphère cérébral lésé etmeilleure est la fonction motrice. Ainsi, la persistanced’activations et de projections controlatérales aumouvement (notamment via le faisceau corticospinalcroisé) est corrélée avec une meilleure fonction motricede la main parétique [34,35].
En imagerie fonctionnelle, les activations cérébralesretrouvées au niveau du cortex lésé après un mouvementde la main parétique (main controlatérale à la lésion) sesituent dans des zones proches de là où devraient sesituer théoriquement les aires motrices s’il n’y avait paseu de lésion [17,36]. Cette adaptation locale del’activation motrice, c’est-à-dire proche des zonescérébrales normalement dédiées à la motricité est,semble-t-il, un facteur de bon pronostic sur le plan del’efficacité motrice. Plus il existe d’activation proche dusillon central du côté de l’hémisphère lésé, et plus lafonction motrice de la main parétique est bonne. End’autres termes, il semble que plus les régions procheslocalement du gyrus précentral sont préservées (doncfonctionnellement encore actives lors des tâchesmotrices en IRMf) et plus la main controlatérale à lalésion est efficiente sur le plan moteur. Ces données sontà rapprocher des données d’analyse structurale de lasubstance grise. En effet, dans le cadre de la diplégiespastique liée à des séquelles périventriculaires de lasubstance blanche, il semble exister une corrélationentre le volume de substance grise dans les gyri pré- etpost-centraux [23] et la performance motrice. Plus ilexiste de substance grise au niveau de S1-M1 et plus lafonction motrice de la main controlatérale est bonne. Onpeut noter ici, toujours sur le plan structural, qu’il existe
un lien entre le degré d’atteinte de la substance blancheet le volume de substance grise sus-jacent [37]. Ainsi,des lésions intéressant uniquement la substance blancheengendrent une diminution concomitante de la sub-stance grise ipsilatérale à ces lésions.
À l’inverse du bon pronostic fonctionnel lié aurespect de l’activité du cortex S1-M1 ipsilésionnel, il aété montré, notamment chez l’adulte après un AVCmais aussi chez l’enfant [38] pendant un mouvement dela main parétique, que des activations supplémentairesau niveau du cortex sensorimoteur non lésé (activationipsilatérale au mouvement) pouvaient être rattachées àun mauvais facteur pronostic moteur. Lorsque l’onutilise le mouvement de la main parétique commeparadigme d’exploration de la motricité-cérébrale, etnotamment le mouvement passif, c’est-à-dire unparadigme relativement indépendant de la performancemotrice, et que l’on compare les cartographiesd’activations cérébrales obtenues chez des enfantssains avec celles obtenues chez des enfants présentantune PC unilatérale, on note chez ces derniers, desactivations plus importantes de l’ensemble du cortexsensorimoteur et ce surtout du côté ipsilatéral aumouvement (du côté du cortex non lésé) [39]. Cetteadaptation, avec des activations du cortex sensorimo-teur non lésé, semble être en fait une « mal-adaptation »,puisque les sujets présentant ce type de plasticité post-lésionnelle ont une mauvaise fonction motrice de leurmain parétique [40]. Aussi cette activation surnumé-raire du cortex sensorimoteur non lésé lors desmouvements de la main parétique semble péjorativepour le fonctionnement moteur de la main parétique.Les explications données quant à la présence de cesactivations ipsilatérales sont mal connues et encorediscutées : persistance du faisceau pyramidal direct ?,défaut d’inhibition motrice ?. . .
Chez l’adulte, après un AVC, la préservation ou nonde la balance inhibitrice inter-hémisphérique joue unrôle important dans la fonction motrice de la mainparétique. Des excès d’inhibition du cortex moteur sainsur le cortex moteur lésé ont été constatés avec unimpact péjoratif sur la fonction motrice de la mainatteinte [41]. Ces évidences ont également été montréeschez des enfants ayant subi un AVC sous-cortical[34,35]. Il nous semble que cette dérégulation de labalance inhibitrice est sous-étudiée chez les enfantsavec PC et que la part de celle-ci dans la fonctionmotrice globale de l’enfant est aussi sous-évaluée.
Il a été montré que le respect de l’intégrité des fibresdu corpus callosum était corrélé à une meilleureperformance dans les activités bi-manuelles chez desenfants avec PC unilatérale [36]. Ces auteurs ont

M. Dinomais et al. / Motricité cérébrale 35 (2014) 3–14 11
souligné l’importance des connexions inter-hémisphéri-ques dans les performances motrices des personnes avecPC. Peut-être s’agit-il, ici, d’une illustration du respectde la balance inhibitrice inter-hémisphérique via lerespect de l’intégrité des fibres inter-hémisphériques ?En effet, les deux hémisphères sont connectés dupoint de vue structurel et fonctionnel grâce auxconnexions trans-callosales, et ces connexions semblentêtre majoritairement inhibitrices (inhibition inter-hémisphérique). Or après une lésion hémisphériqueunilatérale, il peut exister un déséquilibre de la balanceinter-hémisphérique avec une diminution de l’inhibitiondu cortex lésé sur le cortex sain et une augmentation del’inhibition du cortex sain sur le cortex lésé. Comme vuplus haut, chez l’adulte après un AVC unilatéral, cedéséquilibre de la balance inhibitrice inter-hémis-phérique est alors péjoratif pour la fonction motricede la main parétique. La restauration cette balance estun facteur de bon pronostic de récupération motrice[42]. Il en est probablement de même pour l’enfant avecPC unilatérale.
3.3. Plasticité du système sensitif
Quel que soit le type de réorganisation motrice lorsde la PC, il n’existe pas de réorganisation inter-hémisphérique de S1. Ainsi, S1 est toujours situé auniveau controlatéral à la main testée [43,44]. L’absencede réorganisation inter-hémisphérique de S1 est pro-bablement liée au développement et à la mise en placedes voies corticales de la sensibilité qui se déroulent surune longue période à partir du troisième trimestre degrossesse et qui se terminent après la naissance. Ainsien cas de lésion cérébrale précoce, les fibres nerveusessensitives thalamo-corticales contournent cette lésion(même si celle-ci est importante) pour gagner leur zonecible (S1) [45]. Il semble que le système sensitif ne soitpas capable d’une plasticité importante en raison d’unedétermination précoce de la fonction de ces airescérébrales [19].
Nous avons montré [46] que, quel soit le type delésion précoce, le cross-over entre S1 (de l’hémisphèrelésé) et le cervelet est préservé et que S1 se situetoujours au niveau de l’hémisphère lésé quel que soit lemode de réorganisation du système moteur après lésioncérébrale unilatérale précoce. Le système sensitifsemble donc peu doué de plasticité post-lésionnelle.
Il faut noter alors que pour les enfants ayant uneréorganisation ipsilatérale du cortex moteur, il existeune dissociation S1-M1 du contrôle de la mainparétique, avec M1 situé au niveau du gyrus précentralnon lésé, et S1 situé au niveau du gyrus post-central
lésé. L’influence d’une telle dissociation sur lesperformances motrices n’est pas connue. Cette cons-tatation explique peut-être pourquoi la contrainteinduite est moins efficace pour les sujets avecréorganisation motrice ipsilatérale que controlatérale[47]. La question de la répercussion des troublessensitifs sur la fonction motrice reste entière [48].
Hoon et al. [49] ont montré, chez des enfants, nésprématurés, souffrant de PC liée à une atteintepériventriculaire de la substance blanche, que le degréd’atteinte des radiations du faisceau thalamo-cortical,se projetant du thalamus sensitif vers le cortex pariétal(gyrus post-central et pariétal postérieur), était corréléavec les difficultés sensitives de l’enfant mais aussi avecses difficultés motrices. Notons qu’il n’avait pas étéretrouvé, dans le travail de Hoon et al. [49], decorrélation entre le niveau d’atteinte du faisceaupyramidal et le degré de difficultés sensitives, nimême avec le degré de difficulté motrice et celacontrairement à ce que nous avons exposé précédem-ment, notamment par rapport aux données de Lee et al.[23].
3.4. Réseau de l’observation du mouvement chezl’enfant avec PC
Chez le sujet sans lésion cérébrale, l’observationd’un mouvement effectué par un tiers augmentesélectivement l’excitabilité des pools neuronaux asso-ciés à la production de ce mouvement chez l’observa-teur alors même que le sujet n’effectue pas réellement lemouvement. Un des substrats neuronaux à cephénomène est connu sous le terme de « neuronemiroir » [50]. Ces neurones ont pour caractéristiqueprincipale de répondre sur le plan électrophysiologique(décharge neuronale) à la fois lorsque le sujet produitune action dirigée vers un but et lorsque celui-ci observecette même action. Si un neurone ne possède pas cesdeux caractéristiques, il ne peut être considéré commeun « neurone miroir ». Il s’agit donc d’une définitionélectrophysiologique initialement décrite chez leprimate non humain [50]. Cependant, certaines donnéesen imagerie fonctionnelle peuvent suggérer l’existencede tels neurones chez l’homme [51,52].
De façon plus générale, et en s’affranchissant de ladéfinition précise électrophysiologique des neurones àpropriété visuo-motrice (« neurones miroirs » notam-ment), les données d’imagerie fonctionnelle ont permisd’affirmer que le fait d’observer quelqu’un en train deréaliser une action se traduit par une activation« automatique » des réseaux moteurs cérébraux sous-tendant la réalisation de cette action chez l’observateur.

M. Dinomais et al. / Motricité cérébrale 35 (2014) 3–1412
On parle de réseau de l’observation de l’action « actionobservation network ».
De nombreuses études ont validé l’existence d’uneactivation de M1 lors de l’observation du mouvementbiologique qu’il soit dirigé vers un but (mouvement dittransitif) ou non (mouvement intransitif). Stefan et al.[53] ont aussi montré que l’observation répétée d’ungeste simple entraîne une réorganisation du systèmemoteur cortical. Cependant, il a été montré que ce lienexistant entre l’observation d’une action et l’activationdes structures motrices liées à la réalisation de cetteaction (lien visuo-moteur) n’est pas inné, mais acquispar l’expérience [54].
Dans le cadre de la PC unilatérale où le sujetprésente une expérience sensorimotrice faible auniveau de sa main parétique, on peut se poser laquestion de l’existence de telles activations cérébraleslors de l’observation du mouvement. En effet,l’observation d’une action par un enfant avec PCentraîne-t-elle, comme chez le sujet sain, l’activationdes réseaux neuronaux responsables de cette action ?Dans une récente étude [55] chez des enfants avec PCunilatérale ayant une réorganisation controlatérale deM1, nous avons montré que l’observation d’unmouvement simple d’une main parétique entraînel’activation du cortex sensorimoteur notamment ducôté lésé et ce indépendamment du degré de difficultésmotrices de l’enfant. Par ailleurs, nous avons mis enévidence que le caractère croisé de la motricité estconservé lors de l’observation du mouvement. Ainsi, ilexiste un lien visuo-moteur chez l’enfant avec PCunilatérale lors de l’observation d’un mouvementd’ouverture–fermeture de main, et l’observation dela main du côté parétique entraîne une activation plusimportante du cortex lésé que l’observation d’une maindu côté sain.
4. Impact pour les techniques de rééducation
Ces données sur la réorganisation de la motricité-cérébrale nous permettent de confirmer l’intérêt decertaines de nos techniques de rééducation et d’enévoquer d’autres.
Ainsi, le mouvement passif de la main parétique, quisert avant tout à lutter contre les rétractions musculaireset à prévenir les limitations de mobilité articulaire,apparaît comme une tâche rééducative qui stimule chezl’enfant avec PC, le cortex moteur lésé mais aussil’hémisphère controlatéral (hémisphère sain) [56]. Laréalisation de mouvement passif de la main parétiquesemble donc pouvoir stimuler tout ou partie de lamotricité-cérébrale au niveau de l’hémisphère lésé et
ainsi favoriser la plasticité post-lésionnelle en luttantcontre le phénomène de non-utilisation du membreparétique. Il apparaît alors que le mouvement passifpeut, outre son effet de prévention orthopédique, avoirun effet sur la plasticité post-lésionnelle en stimulantl’activité des aires cérébrales périlésionnelles proche dela région centrale ipsilatérale à la lésion. Cettetechnique est particulièrement utile pour les patientsqui ne sont plus capables de réaliser de mouvement actifavec leur main parétique.
L’utilisation contrainte du membre supérieur paré-tique, par l’utilisation de la contrainte induite (tech-nique initialement développée chez l’adulte après AVC)a montré son efficacité clinique chez l’enfant avec PC[57]. Une très récente étude en imagerie a permis demontrer que l’amélioration clinique motrice observéechez les enfants avec PC après une telle rééducationpouvait être mise en lien avec une augmentation duvolume de la substance grise au niveau du cortexsensorimoteur lésé [58]. De notre point de vue, ce faitclinique est à mettre en lien avec l’existence d’unecorrélation positive entre la quantité de substance grisedans M1 lésée et la fonction motrice de la mainparétique. Ces données de neuro-imagerie, en lien avecles améliorations cliniques observées après contrainteinduite, permettent de proposer l’utilisation de cettetechnique dans le cadre de la rééducation de l’enfantavec PC. Cependant et bien que les raisons n’en soientpas bien connues, la contrainte induite paraît ne pasbénéficier également à tous les enfants avec PCunilatérale. En effet, les enfants présentant uneréorganisation ipsilatérale de M1 semblent peut profitersur le plan clinique d’une telle tâche de rééducation[47]. Le type de réorganisation de la motricité-cérébraleinflue donc probablement sur l’efficacité de certainesinterventions rééducatives [59].
En se fondant sur la théorie de l’observation dumouvement et son effet sur l’activation cérébrale desenfants avec PC, il a été testé cliniquement unerééducation à base d’observation du mouvement chezdes enfants avec PC avec des résultats moteursfonctionnels tout à fait encourageants [60–62].
5. Conclusion
Le système sensorimoteur après lésion cérébraleprécoce est donc doué d’une certaine adaptation pourminimiser au mieux les répercussions sur la perte desfonctions sensorimotrices. La réorganisation dusystème moteur peut être intra-hémisphérique (réorga-nisation controlatérale de M1 par exemple après AVCnéonatal) ou inter-hémisphérique et homotopique

M. Dinomais et al. / Motricité cérébrale 35 (2014) 3–14 13
(réorganisation ipsilatérale après lésions périventricu-laires). Le système sensitif semble lui peu plastique, etS1 se situe toujours au niveau de l’hémisphère lésé. Cecia pour conséquence l’existence d’une dissociationS1-M1 en cas de réorganisation ipsilatérale du cortexmoteur. Le type de réorganisation de la motricité-cérébrale semble influer sur l’efficacité de certainestâches rééducatives (notamment la contrainte induite[47]). La MPR doit prendre en compte ces données pourproposer des méthodes de rééducation dans le cadre dela neuro-rééducation des troubles moteurs.
Déclaration d’intérêts
Les auteurs déclarent ne pas avoir de conflitsd’intérêts en relation avec cet article.
Références
[1] Grossiord A. Éditorial. Motricite Cereb 1980;1:1–3.[2] Hertz-Pannier L. Plasticité au cours de la maturation cérébrale :
bases physiologiques et étude par IRM fonctionnelle. J Neuro-radiol 1999;26:66–74.
[3] Faralli A, Bigoni M, Mauro A, Rossi F, Carulli D. Noninvasivestrategies to promote functional recovery after stroke. NeuralPlast 2013;2013:854597.
[4] Kennard M. Age and other factors in motor recovery fromprecentral lesions in monkeys. Am J Physiol 1936;115:138–46.
[5] Christine C, Dolk H, Platt MJ, Colver A, Prasauskiene A,Krageloh-Mann I. Recommendations from the SCPE collabo-rative group for defining and classifying cerebral palsy. Dev MedChild Neurol Suppl 2007;109:35–8.
[6] Rosenbaum P, Paneth N, Leviton A, Goldstein M, Bax M,Damiano D, et al. A report: the definition and classificationof cerebral palsy. Dev Med Child Neurol Suppl 2007;109:8–14.
[7] Stanley F, Blair E, Alberman E, editors. Cerebral palsies:epidemiology and causal pathways in clinics in developmentalmedicine. London: Mac-Keith Press; 2000. p. 151.
[8] Krageloh-Mann I, Cans C. Cerebral palsy update. Brain Dev2009;31(7):537–44.
[9] Eyre JA, Miller S, Clowry GJ, Conway EA, Watts C. Functionalcorticospinal projections are established prenatally in the humanfoetus permitting involvement in the development of spinalmotor centres. Brain 2000;123(Pt 1):51–64.
[10] Eyre JA, Taylor JP, Villagra F, Smith M, Miller S. Evidence ofactivity-dependent withdrawal of corticospinal projections dur-ing human development. Neurology 2001;57(9):1543–54.
[11] Muller K, Kass-Iliyya F, Reitz M. Ontogeny of ipsilateralcorticospinal projections: a developmental study with transcra-nial magnetic stimulation. Ann Neurol 1997;42(5):705–11.
[12] Galea MP, Darian-Smith I. Postnatal maturation of the directcorticospinal projections in the macaque monkey. Cereb Cortex1995;5(6):518–40.
[13] Martin JH, Lee SJ. Activity-dependent competition betweendeveloping corticospinal terminations. Neuroreport 1999;10(11):2277–82.
[14] Martin JH, Kably B, Hacking A. Activity-dependent develop-ment of cortical axon terminations in the spinal cord and brainstem. Exp Brain Res 1999;125(2):184–99.
[15] Olivier E, Edgley SA, Armand J, Lemon RN. An electrophy-siological study of the postnatal development of the corticospinalsystem in the macaque monkey. J Neurosci 1997;17(1):267–76.
[16] Armand J, Olivier E, Edgley SA, Lemon RN. Postnatal deve-lopment of corticospinal projections from motor cortex to thecervical enlargement in the macaque monkey. J Neurosci1997;17(1):251–66.
[17] Staudt M, Grodd W, Gerloff C, Erb M, Stitz J, Krageloh-Mann I.Two types of ipsilateral reorganization in congenital hemipare-sis: a TMS and fMRI study. Brain 2002;125(Pt 10):2222–37.
[18] Kostovic I, Judas M. Correlation between the sequentialingrowth of afferents and transient patterns of cortical laminationin preterm infants. Anat Rec 2002;267(1):1–6.
[19] Juenger H, de Haan B, Krageloh-Mann I, Staudt M, Karnath HO.Early determination of somatosensory cortex in the human brain.Cereb Cortex 2011;21(8):1827–31.
[20] Holmstrom L, Vollmer B, Tedroff K, Islam M, Persson JK, KitsA, et al. Hand function in relation to brain lesions and cortico-motor-projection pattern in children with unilateral cerebralpalsy. Dev Med Child Neurol 2010;52(2):145–52.
[21] Holmefur M, Kits A, Bergstrom J, Krumlinde-Sundholm L,Flodmark O, Forssberg H, et al. Neuroradiology can predictthe development of hand function in children with unilateralcerebral palsy. Neurorehabil Neural Repair 2013;27(1):72–8.
[22] Husson B, Hertz-Pannier L, Renaud C, Allard D, Presles E,Landrieu P, et al. Motor outcomes after neonatal arterial ische-mic stroke related to early MRI data in a prospective study.Pediatrics 2010;126(4):912–8.
[23] Lee JD, Park HJ, Park ES, Oh MK, Park B, Rha DW, et al. Motorpathway injury in patients with periventricular leucomalacia andspastic diplegia. Brain 2011;134(Pt 4):1199–210.
[24] Duque J, Thonnard JL, Vandermeeren Y, Sebire G, Cosnard G,Olivier E. Correlation between impaired dexterity and corticos-pinal tract dysgenesis in congenital hemiplegia. Brain 2003;126(Pt 3):732–47.
[25] Gordon AL, Wood A, Tournier JD, Hunt RW. Corticospinal tractintegrity and motor function following neonatal stroke: a casestudy. BMC Neurol 2012;12:53.
[26] van der Aa NE, Verhage CH, Groenendaal F, Vermeulen RJ, deBode S, van Nieuwenhuizen O, et al. Neonatal neuroimagingpredicts recruitment of contralesional corticospinal tracts followingperinatal brain injury. Dev Med Child Neurol 2013;55(8):707–12.
[27] Eyre JA, Smith M, Dabydeen L, Clowry GJ, Petacchi E, BattiniR, et al. Is hemiplegic cerebral palsy equivalent to amblyopia ofthe corticospinal system? Ann Neurol 2007;62(5):493–503.
[28] Eyre JA. Corticospinal tract development and its plasticity afterperinatal injury. Neurosci Biobehav Rev 2007;31(8):1136–49.
[29] Maegaki Y, Maeoka Y, Ishii S, Shiota M, Takeuchi A, Yoshino K,et al. Mechanisms of central motor reorganization in pediatrichemiplegic patients. Neuropediatrics 1997;28(3):168–74.
[30] Staudt M. Reorganization after pre-, perinatal brain lesions.J Anat 2010;217(4):469–74.
[31] Staudt M, Braun C, Gerloff C, Erb M, Grodd W, Krageloh-MannI. Developing somatosensory projections bypass periventricularbrain lesions. Neurology 2006;67(3):522–5.
[32] Staudt M, Gerloff C, Grodd W, Holthausen H, Niemann G,Krageloh-Mann I. Reorganization in congenital hemiparesisacquired at different gestational ages. Ann Neurol 2004;56(6):854–63.
[33] Gallea C, Popa T, Billot S, Meneret A, Depienne C, Roze E.Congenital mirror movements: a clue to understanding bimanualmotor control. J Neurol 2011;258(11):1911–9.

M. Dinomais et al. / Motricité cérébrale 35 (2014) 3–1414
[34] Kirton A, Deveber G, Gunraj C, Chen R. Cortical excitabilityand interhemispheric inhibition after subcortical pediatricstroke: plastic organization and effects of rTMS. Clin Neuro-physiol 2010;121(11):1922–9.
[35] Kirton A, Chen R, Friefeld S, Gunraj C, Pontigon AM, DeveberG. Contralesional repetitive transcranial magnetic stimulationfor chronic hemiparesis in subcortical paediatric stroke: a ran-domised trial. Lancet Neurol 2008;7(6):507–13.
[36] Weinstein M, Green D, Geva R, Schertz M, Fattal-Valevski A,Artzi M, et al. Interhemispheric and intrahemispheric connectivityand manual skills in children with unilateral cerebral palsy. BrainStruct Funct 2013. http://dx.doi.org/10,1007/s00429-013-0551-5.
[37] Li D, Hodge J, Wei XC, Kirton A. Reduced ipsilesional corticalvolumes in fetal periventricular venous infarction. Stroke2012;43(5):1404–7.
[38] Cao Y, Vikingstad EM, Huttenlocher PR, Towle VL, Levin DN.Functional magnetic resonance studies of the reorganization ofthe human hand sensorimotor area after unilateral brain injury inthe perinatal period. Proc Natl Acad Sci U S A 1994;91(20):9612–6.
[39] Van de Winckel A, Klingels K, Bruyninckx F, Wenderoth N,Peeters R, Sunaert S, et al. How does brain activation differ inchildren with unilateral cerebral palsy compared to typicallydeveloping children, during active and passive movements, andtactile stimulation? An fMRI study. Res Dev Disabil 2012;34(1):183–97.
[40] Alagona G, Delvaux V, Gerard P, De Pasqua V, Pennisi G,Delwaide PJ, et al. Ipsilateral motor responses to focal trans-cranial magnetic stimulation in healthy subjects and acute-strokepatients. Stroke 2001;32(6):1304–9.
[41] Murase N, Duque J, Mazzocchio R, Cohen LG. Influence ofinterhemispheric interactions on motor function in chronicstroke. Ann Neurol 2004;55(3):400–9.
[42] Hummel FC, Cohen LG. Non-invasive brain stimulation: a newstrategy to improve neurorehabilitation after stroke? LancetNeurol 2006;5(8):708–12.
[43] Wilke M, Staudt M, Juenger H, Grodd W, Braun C, Krageloh-Mann I. Somatosensory system in two types of motor reorga-nization in congenital hemiparesis: topography and function.Hum Brain Mapp 2009;30(3):776–88.
[44] Guzzetta A, Bonanni P, Biagi L, Tosetti M, Montanaro D, GuerriniR, et al. Reorganisation of the somatosensory system after earlybrain damage. Clin Neurophysiol 2007;118(5):1110–21.
[45] Staudt M, Erb M, Braun C, Gerloff C, Grodd W, Krageloh-MannI. Extensive peri-lesional connectivity in congenital hemiparesis.Neurology 2006;66(5):771.
[46] Dinomais M, Groeschel S, Staudt M, Krageloh-Mann I, WilkeM. Relationship between functional connectivity and sensoryimpairment: red flag or red herring? Hum Brain Mapp 2012;33(3):628–38.
[47] Kuhnke N, Juenger H, Walther M, Berweck S, Mall V, Staudt M.Do patients with congenital hemiparesis and ipsilateral corti-cospinal projections respond differently to constraint-inducedmovement therapy? Dev Med Child Neurol 2008;50(12):898–903.
[48] Wilke M, Staudt M. Does damage to somatosensory circuitsunderlie motor impairment in cerebral palsy? Dev Med ChildNeurol 2009;51(9):686–7.
[49] Hoon Jr AH, Stashinko EE, Nagae LM, Lin DD, Keller J, BastianA, et al. Sensory and motor deficits in children with cerebralpalsy born preterm correlate with diffusion tensor imagingabnormalities in thalamocortical pathways. Dev Med ChildNeurol 2009;51(9):697–704.
[50] Gallese V, Fadiga L, Fogassi L, Rizzolatti G. Action recognitionin the premotor cortex. Brain 1996;119(Pt 2):593–609.
[51] Cattaneo L, Rizzolatti G. The mirror neuron system. ArchNeurol 2009;66(5):557–60.
[52] Rizzolatti G, Craighero L. The mirror-neuron system. Annu RevNeurosci 2004;27:169–92.
[53] Stefan K, Cohen LG, Duque J, Mazzocchio R, Celnik P, SawakiL, et al. Formation of a motor memory by action observation. JNeurosci 2005;25(41):9339–46.
[54] Calvo-Merino B, Glaser DE, Grezes J, Passingham RE, HaggardP. Action observation and acquired motor skills: an FMRI studywith expert dancers. Cereb Cortex 2005;15(8):1243–9.
[55] Dinomais M, Lignon G, Chinier E, Richard I, Ter Minassian A,Tich SN. Effect of observation of simple hand movement onbrain activations in patients with unilateral cerebral palsy: anfMRI study. Res Dev Disabil 2013;34(6):1928–37.
[56] Van de Winckel A, Klingels K, Bruyninckx F, Wenderoth N,Peeters R, Sunaert S, et al. How does brain activation differ inchildren with unilateral cerebral palsy compared to typicallydeveloping children, during active and passive movements, andtactile stimulation? An fMRI study. Res Dev Disabil 2013;34(1):183–97.
[57] Eliasson AC, Krumlinde-sundholm L, Shaw K, Wang C. Effectsof constraint-induced movement therapy in young children withhemiplegic cerebral palsy: an adapted model. Dev Med ChildNeurol 2005;47(4):266–75.
[58] Sterling C, Taub E, Davis D, Rickards T, Gauthier LV, Griffin A,et al. Structural neuroplastic change after constraint-inducedmovement therapy in children with cerebral palsy. Pediatrics2013;131(5):e1664–9.
[59] Juenger H, Kuhnke N, Braun C, Ummenhofer F, Wilke M,Walther M, et al. Two types of exercise-induced neuroplasticityin congenital hemiparesis: a transcranial magnetic stimulation,functional MRI, and magnetoencephalography study. Dev MedChild Neurol 2013;55(10):941–51.
[60] Sgandurra G, Ferrari A, Cossu G, Guzzetta A, Fogassi L, CioniG. Randomized trial of observation and execution of upperextremity actions versus action alone in children with unilateralcerebral palsy. Neurorehabil Neural Repair 2013;27(9):808–15.
[61] Buccino G, Arisi D, Gough P, Aprile D, Ferri C, Serotti L, et al.Improving upper limb motor functions through action observa-tion treatment: a pilot study in children with cerebral palsy. DevMed Child Neurol 2012;54(9):822–8.
[62] Sgandurra G, Ferrari A, Cossu G, Guzzetta A, Biagi L, Tosetti M,et al. Upper limb children action-observation training (UP-CAT):a randomised controlled trial in hemiplegic cerebral palsy. BMCNeurol 2011;11:80.