Embed Size (px)
Citation preview

We identifi ed two phenylquinazoline compounds in a large-scale screening for cytokinin antagonists in yeast expressing the Arabidopsis cytokinin receptor cytokinin response 1/histidine kinase 4 (CRE1). After chemical modifi cations, we obtained compound S-4893, which non-competitively inhibited binding of the natural ligand 2-isopentenyladenine to CRE1. S-4893 antagonized cytokinin-induced activation of the Arabidopsis response regulator 5 promoter in Arabidopsis. Importantly, S-4893 had no detectable intrinsic cytokinin agonist activity in Arabidopsis or in the transformed yeast system. Cytokinin bioassay further demonstrated that S-4893 antagonized cytokinin-induced stimulation of callus formation and inhibition of root elongation. S-4893 also promoted seminal, crown and lateral root growth in rice, suggesting that S-4893 could potentially promote root growth in a variety of agronomically important plants. We believe S-4893 will be a useful tool in functional studies of cytokinin action in a wide range of plants and a lead compound for the development of useful root growth promoters in agriculture.
Keywords: Arabidopsis thaliana • Cytokinin • Cytokinin antagonist • Cytokinin receptor • Plant growth regulator • Root growth promoter .
Abbreviations : AHK4 , Arabidopsis histidine kinase 4 ; ARR5 , Arabidopsis response regulator 5 ; BA , N 6 -benzyladenine ; CRE1 , cytokinin response 1 ; DMSO , dimethylsulfoxide ; GUS , β -glucuronidase ; IC 50 , the 50 % inhibitory concentration (concentration that inhibits growth by 50 % ) ; iP , 2-isopentenyladenine ; MOA , mechanism of action ; PGR , plant growth regulator ; RGP , root growth promoter.
Introduction
Substances useful for regulation of plant growth and develop-ment, known as ‘plant growth regulators’ (PGRs), have signifi -cantly contributed to agricultural improvements in the yield and quality of various crops. Several classes of PGRs are com-mercially available for practical application ( Koshioka et al. 2005 ). Stable effi cacy over a broad spectrum of crops is a desirable feature of commercialized PGR products, on the basis of rather small markets in each crop. Plant hormones are common regulators of growth and development in all higher plants. Consequently, many commercialized PGR products are natural or synthetic plant hormones such as auxin, gibberellin, ethylene, cytokinin and ABA, or chemicals with a direct infl u-ence on the biosynthesis or metabolism of endogenous gibber-ellin or ethylene ( Ferguson et al. 2006 ). In commercialized PGR products, this type of clear mechanism of action (MOA) through plant hormones is greatly advantageous to both effi cacy and registrability as a safer agrochemical, and also a priority in research and development of new commercialized PGR products.
Potential market demand for substances suitable for improvement of plant root growth, denoted ‘root growth promoters’ (RGPs), is currently unmet by commercialized PGR products. Root tissue is responsible for crucial functions in plant growth and development, e.g. absorption of nutrients and water from soil, localization of storage substances, anchoring the plant to the ground and supporting the aerial part of plants. However, no RGP with stable effi cacy on root growth and a known MOA has been developed to date. This situation motivated us to develop new RGPs characterized by a con-fi rmed MOA relevant to plant hormones.
The Phenylquinazoline Compound S-4893 is a Non-Competitive Cytokinin Antagonist that Targets Arabidopsis Cytokinin Receptor CRE1 and Promotes Root Growth in Arabidopsis and Rice Yuto Arata 1 , * , Asako Nagasawa-Iida 1 , Hideki Uneme 1 , Hiroki Nakajima 1 , Tatsuo Kakimoto 2 and Ryo Sato 3 1 Agricultural Chemicals Research Laboratory, Sumitomo Chemical Company Limited, Takarazuka, Hyogo, 665-8555 Japan 2 Department of Biology, Graduate School of Science, Osaka University, Toyonaka, Osaka, 560-0043 Japan 3 Planning and Coordination Offi ce, Agricultural Chemicals Sector, Sumitomo Chemical Company Limited, Chuo-ku, Tokyo, 104-8260 Japan *Corresponding author: E-mail, [email protected] ; Fax, + 81-797-74-2133 (Received July 7, 2010; Accepted October 25, 2010)
Plant Cell Physiol. 51(12): 2047–2059 (2010) doi:10.1093/pcp/pcq163, available FREE online at www.pcp.oxfordjournals.org© The Author 2010. Published by Oxford University Press on behalf of Japanese Society of Plant Physiologists.All rights reserved. For permissions, please email: [email protected]
Editor-in-Chief’s choice
2047Plant Cell Physiol. 51(12): 2047–2059 (2010) doi:10.1093/pcp/pcq163 © The Author 2010.
Regular PaperD
ownloaded from
https://academic.oup.com
/pcp/article/51/12/2047/1944222 by guest on 05 July 2022

Cytokinins, which are a class of plant hormones, centrally regulate cell division and differentiation in plants. They are chemically classifi ed as either adenine derivatives such as zeatin, kinetin and N 6 -benzyladenine (BA), or phenylurea derivatives including diphenylurea and thidiazuron. Since the discovery of kinetin in the 1950s, a variety of studies have demonstrated that cytokinins are related to root development. Cytokinins inhibit formation of adventitious roots and lateral roots as well as root elongation ( Nishimura et al. 2004 , Kuroha et al. 2006 , Li et al. 2006 , Dello Ioio et al. 2007 , Kyozuka 2007 , Benková and Hejátko 2009 ). In contrast, enhanced root development was demonstrated in transgenic tobacco ( Werner et al. 2001 ) and Arabidopsis ( Werner et al, 2003 ) that overexpressed a gene encoding cytokinin oxidase/dehydrogenase, which degrades cytokinins. Additionally, multiple mutants for genes encoding cytokinin biosynthetic enzymes were reported to show enhanced root development ( Miyawaki et al. 2006 ). Plants respond to cytokinins through a multistep phosphorelay system, and cytokinin receptors, which are histidine kinases, initiate the phosphorelay in response to cytokinins. In Arabi-dopsis, there are three cytokinin receptors, cytokinin response 1, also known as Arabidopsis histidine kinase 4 (CRE1/AHK4), Arabidopsis histidine kinase 2 (AHK2) and Arabidopsis histi-dine kinase 3 (AHK3). Single and double knock-out mutants for one or two Arabidopsis cytokinin receptor genes showed a reduced sensitivity to cytokinin and, somewhat paradoxically, triple mutants for all three cytokinin receptor genes exhibited severe root growth defects with abnormal root vasculature ( Higuchi et al. 2004 , Riefl er et al. 2006 ). Therefore, severe inhibi-tion of cytokinin signaling causes retarded growth, but moder-ate inhibition of cytokinin action promotes root elongation. Thus, cytokinin action in root growth may be a potential MOA for new RGPs.
Anticytokinins are compounds characterized by physiologi-cal inhibition of cytokinin action. Anticytokinins were initially identifi ed by screening for chemicals that inhibited cytokinin action in induced callus formation of tobacco ( Hecht et al. 1971 ). Various classes of anticytokinins that inhibit cytokinin action in callus growth have been reported to date ( Iwamura et al. 1985 , Iwamura 1994 ), but RGP activity has not been observed in these anticytokinins. Because many anticytokinins were identifi ed by screening chemicals structurally related to cytokinins, some anticytokinins also exhibit cytokinin agonist activity ( Iwamura et al. 1983 , Iwamura et al. 1985 , Karanov et al. 1993 ). On the basis of this structural similarity between cytokinins and anticytokinins, a putative MOA of anticytoki-nins was thought to be competitive interaction with cytokinin receptors. However, this putative MOA does not agree with the results of Spíchal et al. (2007) , in which anticytokinins did not interact with cytokinin receptors but inhibited cyclin-dependent kinases.
Spíchal et al. (2009) identifi ed the purine derivative com-pound PI-55, which antagonizes cytokinin activity at the recep-tor level. PI-55 is structurally closely related to the cytokinin BA, but substitutions at specifi c positions on the aromatic side
chain strongly diminish its cytokinin activity and confer antago-nistic properties. PI-55 competitively inhibited the binding of the natural ligand trans -zeatin to the Arabidopsis cytokinin receptor CRE1. However, PI-55 still showed weak cytokinin agonistic activity resulting from partial agonistic interference with the other two Arabidopsis cytokinin receptors, AHK2 and AHK3. Recently, Nisler et al. (2010) found that the new purine derivative compound LGR-991 inhibited CRE1/AHK4 at the receptor level to the same extent as PI-55, and also antagonized AHK3. Furthermore, LGR-991 showed reduced agonistic effects in comparison with PI-55 in the cytokinin-inducible reporter gene assay and in classical cytokinin bioassays.
In recent years, genes involved in detection, biosynthesis and signal transduction of cytokinins have been identifi ed ( Kakimoto 2003 , Ferreira and Kieber 2005 , Ding and Ma 2006 , Hwang and Sakakibara 2006 , Choi and Hwang 2007 ), offering us an opportunity to screen for chemicals that affect cytokinin receptors, biosynthesis or signal transduction in a high-through-put and target-based manner. As an example of target-based bioassays, Inoue et al. (2001) reported that transformed yeast cells ( ∆ sln1 [ CRE1 ]) which lacked the essential histidine kinase gene Sln1 and expressed the Arabidopsis CRE1 gene depended on cytokinins for growth, indicating that cytokinin receptor agonists or antagonists can be theoretically selected by using transformed yeast in a high-throughput manner ( Kakimoto et al. 2002 ). In another study, a transformed yeast strain co-expressing Arabidopsis CRE1 and a cytokinin biosynthesis gene, Arabidopsis thaliana adenylate isopentenyltransferase 4 ( AtIPT4 ) or A. thaliana adenylate isopentenyltransferase 5 ( AtIPT5 ) ( Kakimoto 2001 , Takei et al. 2001 ), in the ∆ sln1 mutant was also created, in which self-produced cytokinins activate the cytokinin receptor to induce histidine kinase activity, which in turn replaces the need for the histidine kinase activity of SLN1 for growth ( Kakimoto and Higuchi 2005 ). This yeast line is potentially applicable for screening of chemicals that target cytokinin biosynthetic enzymes ( Kakimoto and Higuchi 2005 ). Membrane fractions containing a recombinant cytoki-nin receptor, and even living bacteria expressing a cytokinin receptor gene, are useful for cytokinin-binding assays ( Romanov et al. 2005 , Romanov et al. 2006 ). Furthermore, Arabidopsis lines expressing a chimeric gene comprised of a cytokinin-inducible promoter and a reporter gene are also available to test the effect of compounds on cytokinin signal transduction in Arabidopsis ( D'Agostino et al. 2000 ).
In the present study, we screened a large number of com-pounds in a target-based assay for cytokinin receptors and a bioassay for RGPs. We have identifi ed compound S-4893, possessing the chemical structure of a 4-phenylquinazoline, as a potential candidate RGP. We have demonstrated that S-4893 acts as a cytokinin antagonist and binds to CRE1 non-competitively in an in vitro binding assay and has antago-nistic effects on various cytokinin-regulated physiological responses in Arabidopsis. S-4893 promotes root growth in both Arabidopsis and rice, indicating that it could be a lead
2048
Y. Arata et al.
Plant Cell Physiol. 51(12): 2047–2059 (2010) doi:10.1093/pcp/pcq163 © The Author 2010.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/51/12/2047/1944222 by guest on 05 July 2022
compound for the development of useful PGRs that improve root system architecture in agronomically important crops.
Results
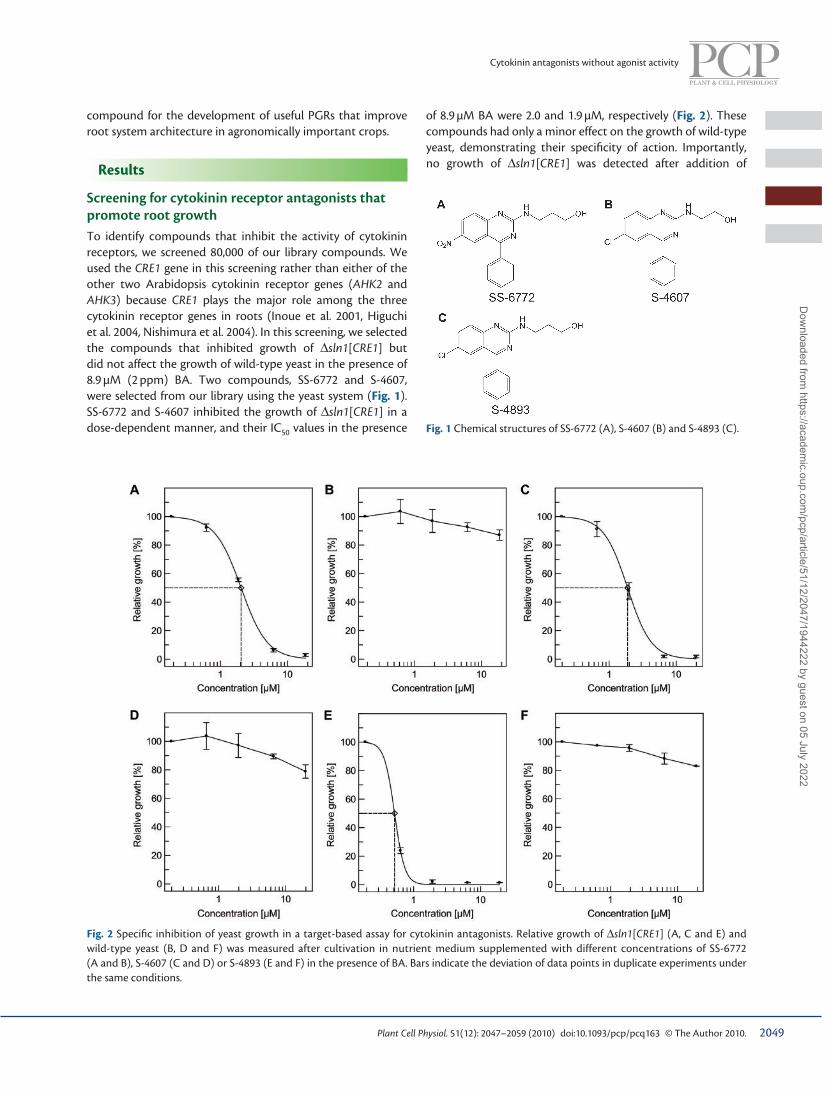
Screening for cytokinin receptor antagonists that promote root growth To identify compounds that inhibit the activity of cytokinin receptors, we screened 80,000 of our library compounds. We used the CRE1 gene in this screening rather than either of the other two Arabidopsis cytokinin receptor genes ( AHK2 and AHK3 ) because CRE1 plays the major role among the three cytokinin receptor genes in roots ( Inoue et al. 2001 , Higuchi et al. 2004 , Nishimura et al. 2004 ). In this screening, we selected the compounds that inhibited growth of ∆ sln1 [ CRE1 ] but did not affect the growth of wild-type yeast in the presence of 8.9 µM (2 ppm) BA. Two compounds, SS-6772 and S-4607, were selected from our library using the yeast system ( Fig. 1 ). SS-6772 and S-4607 inhibited the growth of ∆ sln1 [ CRE1 ] in a dose-dependent manner, and their IC 50 values in the presence Fig. 1 Chemical structures of SS-6772 (A), S-4607 (B) and S-4893 (C).
Fig. 2 Specifi c inhibition of yeast growth in a target-based assay for cytokinin antagonists. Relative growth of ∆ sln1 [ CRE1 ] (A, C and E) and wild-type yeast (B, D and F) was measured after cultivation in nutrient medium supplemented with different concentrations of SS-6772 (A and B), S-4607 (C and D) or S-4893 (E and F) in the presence of BA. Bars indicate the deviation of data points in duplicate experiments under the same conditions.
of 8.9 µM BA were 2.0 and 1.9 µM, respectively ( Fig. 2 ). These compounds had only a minor effect on the growth of wild-type yeast, demonstrating their specifi city of action. Importantly, no growth of ∆ sln1 [ CRE1 ] was detected after addition of
2049
Cytokinin antagonists without agonist activity
Plant Cell Physiol. 51(12): 2047–2059 (2010) doi:10.1093/pcp/pcq163 © The Author 2010.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/51/12/2047/1944222 by guest on 05 July 2022

SS-6772 or S-4607 in the absence of BA ( Table 1 ), demonstrat-ing that these compounds exhibited no detectable agonist activity in this system. Interestingly, SS-6772 and S-4607 shared a common chemical structure of 4-phenylquinazoline, which is structurally distinct from the cytokinin receptor antagonist PI-55 ( Spíchal et al. 2009 ), LGR-991 ( Nisler et al. 2010 ) and all other known anticytokinins or cytokinins ( Fig. 3 ).
A number of compounds have been reported to inhibit the effects of cytokinin on plants ( Hecht et al. 1971 , Iwamura et al. 1979 , Iwamura et al. 1985 , Iwamura 1994 ). These compounds were also subjected to the yeast system. We chose the following fi ve compounds from diverse chemical classes ( Fig. 3 ): pyrido[2,3- d ]pyrimidine (AntiCK-1), triazolo[4,5- d ]pyrimidine (AntiCK-2), benzylphenylurea (AntiCK-3), N -arylcarbamate (AntiCK-4) and substituted pyrazole (AntiCK-5) ( Kefford et al. 1968 , Iwamura et al. 1979 , Karanov et al. 1993 , Iwamura 1994 ). None of these fi ve compounds differentially inhibited the cytokinin-dependent growth of ∆ sln1 [ CRE1 ], suggesting that these compounds do not target cytokinin receptors ( Fig. 4 ).
Because cytokinin antagonists may also increase root growth, we separately screened approximately 40,000 compounds from our library to identify those that enhanced Arabidopsis root growth. The library included a set of compounds that overlapped with that of the library used for selection of cytoki-nin receptor-inhibiting compounds. A 20 % increase in primary root length was set as the threshold of primary selection for signifi cant root elongation after treatment with 2 or 10 ppm test compounds. In this primary screening, 1,818 compounds were chosen, with a primary hit rate of 4.79 % (data not shown). As expected, SS-6772 and S-4607 were included among the positive chemicals identifi ed in the RGP screening, indicating that these compounds may function as RGPs with specifi c actions as cytokinin receptor antagonists.
Fig. 3 Chemical structures of fi ve compounds that have been reported to inhibit cytokinin responses in plants. (1) AntiCK-1, 4-benzylamino-2-methylthiopyrido[2,3- d ]pyrimidine ( Iwamura et al. 1979 ); (2) AntiCK-2, 3-benzyl-7-hydroxy-5-methyl-3 H -1,2,3-triazolo[4,5- d ]pyrimidine ( Karanov et al. 1993 ); (3) AntiCK-3, 1-benzyl-3-(4-chlorophenyl)urea ( Kefford et al. 1968 ); (4) AntiCK-4, propyl N -(3,4-dichlorophenyl)carbamate ( Iwamura 1994 ); and (5) AntiCK-5, ethyl 5-amino-1-benzylpyrazole-4-carboxylate ( Karanov et al. 1993 ).
Table 1 No detectable agonist activity of SS-6772 and S-4607
Compound Concentration (µM)
Average optical density of the wells measured at 600 nm ( n = 12)
Solvent control – 0.036 ± 0.00072
SS-6772 0.19 0.035 ± 0.00072
0.62 0.036 ± 0.00080
1.9 0.036 ± 0.0010
6.2 0.035 ± 0.00094
19 0.035 ± 0.00075
S-4607 0.20 0.035 ± 0.00089
0.66 0.036 ± 0.00065
2.0 0.036 ± 0.0010
6.6 0.035 ± 0.0010
20 0.035 ± 0.00065
No growth of ∆ sln1 [ CRE1 ] was detected after addition of SS-6772 or S-4607 in the absence of BA. Values indicate the average of the optical density of the wells ± SD of 12 independent wells under the same conditions.
To develop compounds that have increased cytokinin antag-onist and root growth-promoting activity, and taking into account the common chemical structure between SS-6772 and S-4607, quinazoline ring substituents at positions 2 and 6 were optimized based on antagonistic activity on transformed yeast and root growth promotion in Arabidopsis. We found that S-4893 demonstrated the strongest inhibition of cytokinin signal transduction in transformed yeast ( Figs. 1 , 2 ). The IC 50 of S-4893 in the yeast system was 0.50 µM, lower than those of SS-6772 and S-4607 ( Fig. 2 ). The root growth-promoting activity of S-4893 was also higher than those of SS-6772 and S-4607 ( Table 2 ) in the bioassay for RGPs.
S-4893 is a non-competitive inhibitor of CRE1 The ability of S-4607 and S-4893 to inhibit binding of 3 H-labeled N -6-(isopentyl-2-enyl)adenine (iP) to the cytokinin receptor CRE1 was examined in an in vitro assay with isolated membrane fractions of ∆ sln1 [ CRE1 ]. Specifi c inhibition by each compound was detected in the assay, albeit to a lesser degree compared with trans -zeatin ( Fig. 5 ). We also evaluated the effect of previ-ously reported anticytokinins in this in vitro assay, but none was effective ( Fig. 5 ). This fi nding is in agreement with the result of the in vivo yeast cytokinin detection system ( Fig. 4 ).
To dissect the mode of inhibition by S-4893, we carried out kinetic analyses. Ligand binding saturation curves at different concentrations of S-4893 or trans -zeatin are illustrated in Fig. 6 . The saturation level of ligand binding was considerably reduced by S-4893 ( Fig. 6A ). This reduction of the saturation level clearly indicated that S-4893 is not a competitive inhibitor of ligand
2050
Y. Arata et al.
Plant Cell Physiol. 51(12): 2047–2059 (2010) doi:10.1093/pcp/pcq163 © The Author 2010.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/51/12/2047/1944222 by guest on 05 July 2022
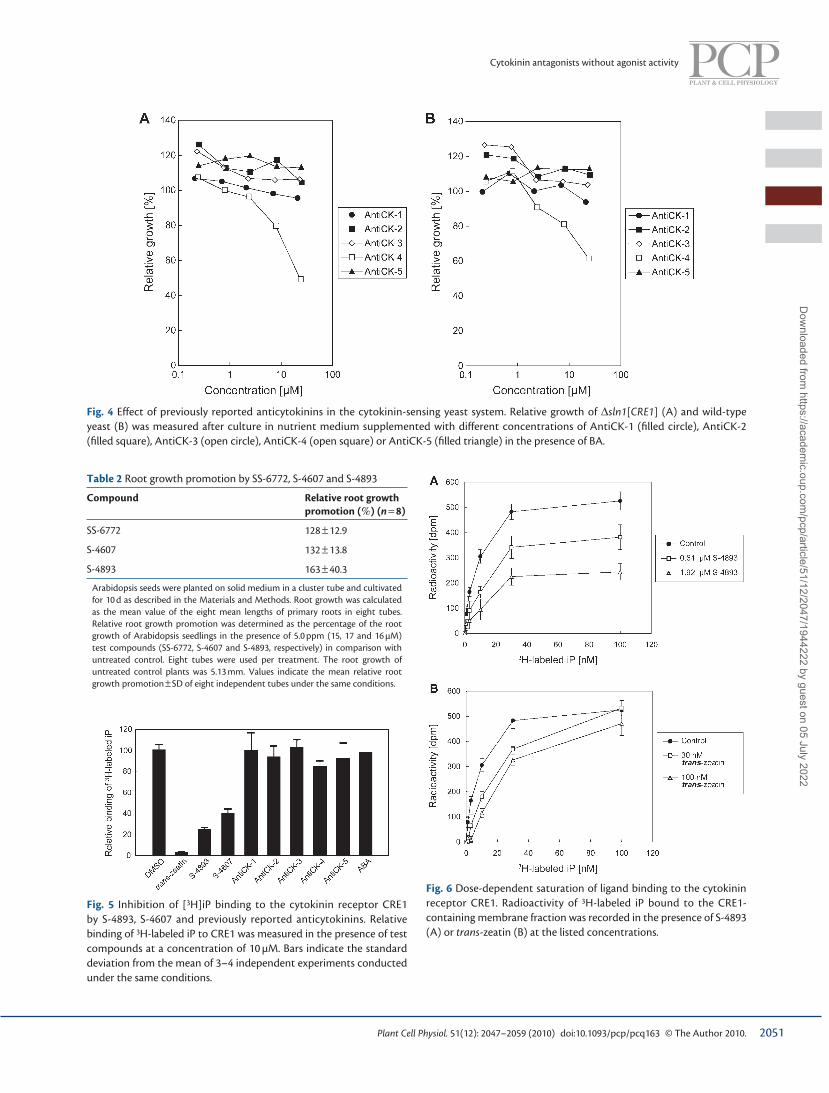
Fig. 4 Effect of previously reported anticytokinins in the cytokinin-sensing yeast system. Relative growth of ∆ sln1 [ CRE1 ] (A) and wild-type yeast (B) was measured after culture in nutrient medium supplemented with different concentrations of AntiCK-1 (fi lled circle), AntiCK-2 (fi lled square), AntiCK-3 (open circle), AntiCK-4 (open square) or AntiCK-5 (fi lled triangle) in the presence of BA.
Fig. 5 Inhibition of [ 3 H]iP binding to the cytokinin receptor CRE1 by S-4893, S-4607 and previously reported anticytokinins. Relative binding of 3 H-labeled iP to CRE1 was measured in the presence of test compounds at a concentration of 10 µM. Bars indicate the standard deviation from the mean of 3–4 independent experiments conducted under the same conditions.
Fig. 6 Dose-dependent saturation of ligand binding to the cytokinin receptor CRE1. Radioactivity of 3 H-labeled iP bound to the CRE1-containing membrane fraction was recorded in the presence of S-4893 (A) or trans -zeatin (B) at the listed concentrations.
Table 2 Root growth promotion by SS-6772, S-4607 and S-4893
Compound Relative root growth promotion ( % ) ( n = 8)
SS-6772 128 ± 12.9
S-4607 132 ± 13.8
S-4893 163 ± 40.3
Arabidopsis seeds were planted on solid medium in a cluster tube and cultivated for 10 d as described in the Materials and Methods. Root growth was calculated as the mean value of the eight mean lengths of primary roots in eight tubes. Relative root growth promotion was determined as the percentage of the root growth of Arabidopsis seedlings in the presence of 5.0 ppm (15, 17 and 16 µM) test compounds (SS-6772, S-4607 and S-4893, respectively) in comparison with untreated control. Eight tubes were used per treatment. The root growth of untreated control plants was 5.13 mm. Values indicate the mean relative root growth promotion ± SD of eight independent tubes under the same conditions.
2051
Cytokinin antagonists without agonist activity
Plant Cell Physiol. 51(12): 2047–2059 (2010) doi:10.1093/pcp/pcq163 © The Author 2010.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/51/12/2047/1944222 by guest on 05 July 2022
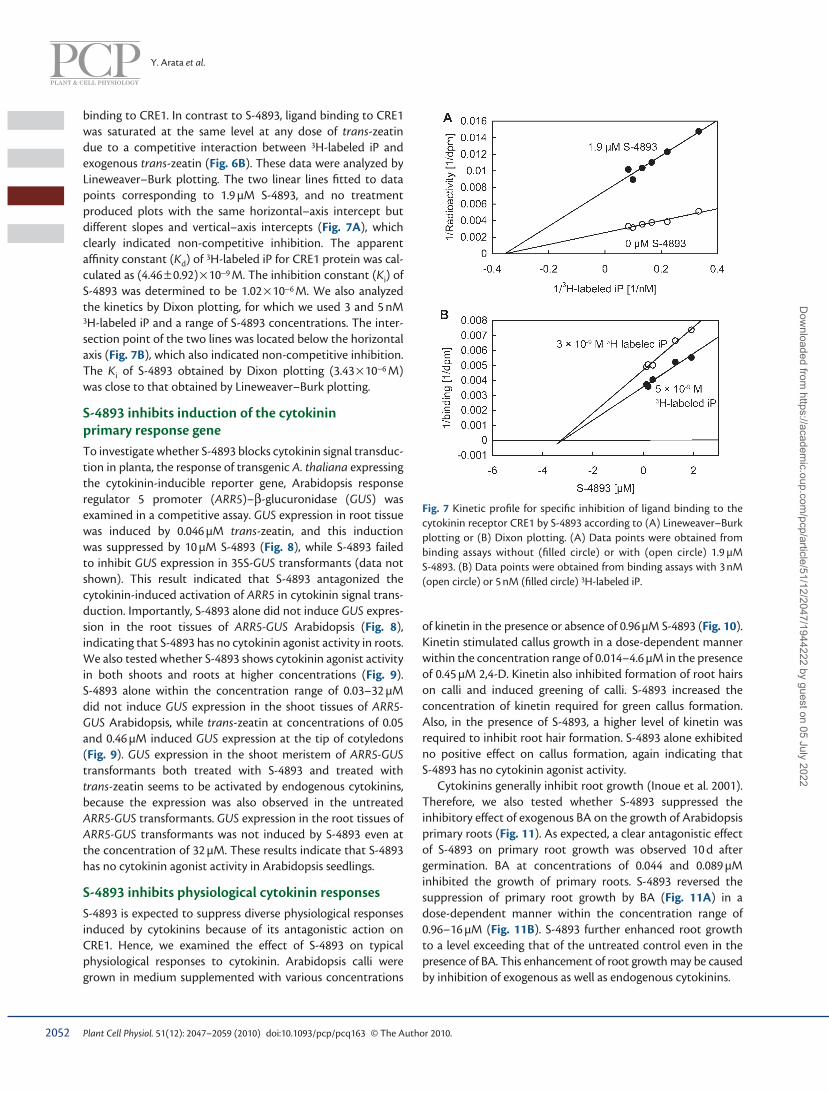
binding to CRE1. In contrast to S-4893, ligand binding to CRE1 was saturated at the same level at any dose of trans -zeatin due to a competitive interaction between 3 H-labeled iP and exogenous trans -zeatin ( Fig. 6B ). These data were analyzed by Lineweaver–Burk plotting. The two linear lines fi tted to data points corresponding to 1.9 µM S-4893, and no treatment produced plots with the same horizontal–axis intercept but different slopes and vertical–axis intercepts ( Fig. 7A ), which clearly indicated non-competitive inhibition. The apparent affi nity constant ( K d ) of 3 H-labeled iP for CRE1 protein was cal-culated as (4.46 ± 0.92) × 10 − 9 M. The inhibition constant ( K i ) of S-4893 was determined to be 1.02 × 10 − 6 M. We also analyzed the kinetics by Dixon plotting, for which we used 3 and 5 nM 3 H-labeled iP and a range of S-4893 concentrations. The inter-section point of the two lines was located below the horizontal axis ( Fig. 7B ), which also indicated non-competitive inhibition. The K i of S-4893 obtained by Dixon plotting (3.43 × 10 − 6 M) was close to that obtained by Lineweaver–Burk plotting.
S-4893 inhibits induction of the cytokinin primary response gene To investigate whether S-4893 blocks cytokinin signal transduc-tion in planta, the response of transgenic A. thaliana expressing the cytokinin-inducible reporter gene, Arabidopsis response regulator 5 promoter ( ARR5 )– β -glucuronidase ( GUS ) was examined in a competitive assay. GUS expression in root tissue was induced by 0.046 µM trans -zeatin, and this induction was suppressed by 10 µM S-4893 ( Fig. 8 ), while S-4893 failed to inhibit GUS expression in 35S- GUS transformants (data not shown). This result indicated that S-4893 antagonized the cytokinin-induced activation of ARR5 in cytokinin signal trans-duction. Importantly, S-4893 alone did not induce GUS expres-sion in the root tissues of ARR5-GUS Arabidopsis ( Fig. 8 ), indicating that S-4893 has no cytokinin agonist activity in roots. We also tested whether S-4893 shows cytokinin agonist activity in both shoots and roots at higher concentrations ( Fig. 9 ). S-4893 alone within the concentration range of 0.03–32 µM did not induce GUS expression in the shoot tissues of ARR5-
GUS Arabidopsis, while trans -zeatin at concentrations of 0.05 and 0.46 µM induced GUS expression at the tip of cotyledons ( Fig. 9 ). GUS expression in the shoot meristem of ARR5-GUS transformants both treated with S-4893 and treated with trans -zeatin seems to be activated by endogenous cytokinins, because the expression was also observed in the untreated ARR5-GUS transformants. GUS expression in the root tissues of ARR5-GUS transformants was not induced by S-4893 even at the concentration of 32 µM. These results indicate that S-4893 has no cytokinin agonist activity in Arabidopsis seedlings.
S-4893 inhibits physiological cytokinin responses S-4893 is expected to suppress diverse physiological responses induced by cytokinins because of its antagonistic action on CRE1. Hence, we examined the effect of S-4893 on typical physiological responses to cytokinin. Arabidopsis calli were grown in medium supplemented with various concentrations
Fig. 7 Kinetic profi le for specifi c inhibition of ligand binding to the cytokinin receptor CRE1 by S-4893 according to (A) Lineweaver–Burk plotting or (B) Dixon plotting. (A) Data points were obtained from binding assays without (fi lled circle) or with (open circle) 1.9 µM S-4893. (B) Data points were obtained from binding assays with 3 nM (open circle) or 5 nM (fi lled circle) 3 H-labeled iP.
of kinetin in the presence or absence of 0.96 µM S-4893 ( Fig. 10 ). Kinetin stimulated callus growth in a dose-dependent manner within the concentration range of 0.014–4.6 µM in the presence of 0.45 µM 2,4-D. Kinetin also inhibited formation of root hairs on calli and induced greening of calli. S-4893 increased the concentration of kinetin required for green callus formation. Also, in the presence of S-4893, a higher level of kinetin was required to inhibit root hair formation. S-4893 alone exhibited no positive effect on callus formation, again indicating that S-4893 has no cytokinin agonist activity.
Cytokinins generally inhibit root growth ( Inoue et al. 2001 ). Therefore, we also tested whether S-4893 suppressed the inhibitory effect of exogenous BA on the growth of Arabidopsis primary roots ( Fig. 11 ). As expected, a clear antagonistic effect of S-4893 on primary root growth was observed 10 d after germination. BA at concentrations of 0.044 and 0.089 µM inhibited the growth of primary roots. S-4893 reversed the suppression of primary root growth by BA ( Fig. 11A ) in a dose-dependent manner within the concentration range of 0.96–16 µM ( Fig. 11B ). S-4893 further enhanced root growth to a level exceeding that of the untreated control even in the presence of BA. This enhancement of root growth may be caused by inhibition of exogenous as well as endogenous cytokinins.
2052
Y. Arata et al.
Plant Cell Physiol. 51(12): 2047–2059 (2010) doi:10.1093/pcp/pcq163 © The Author 2010.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/51/12/2047/1944222 by guest on 05 July 2022
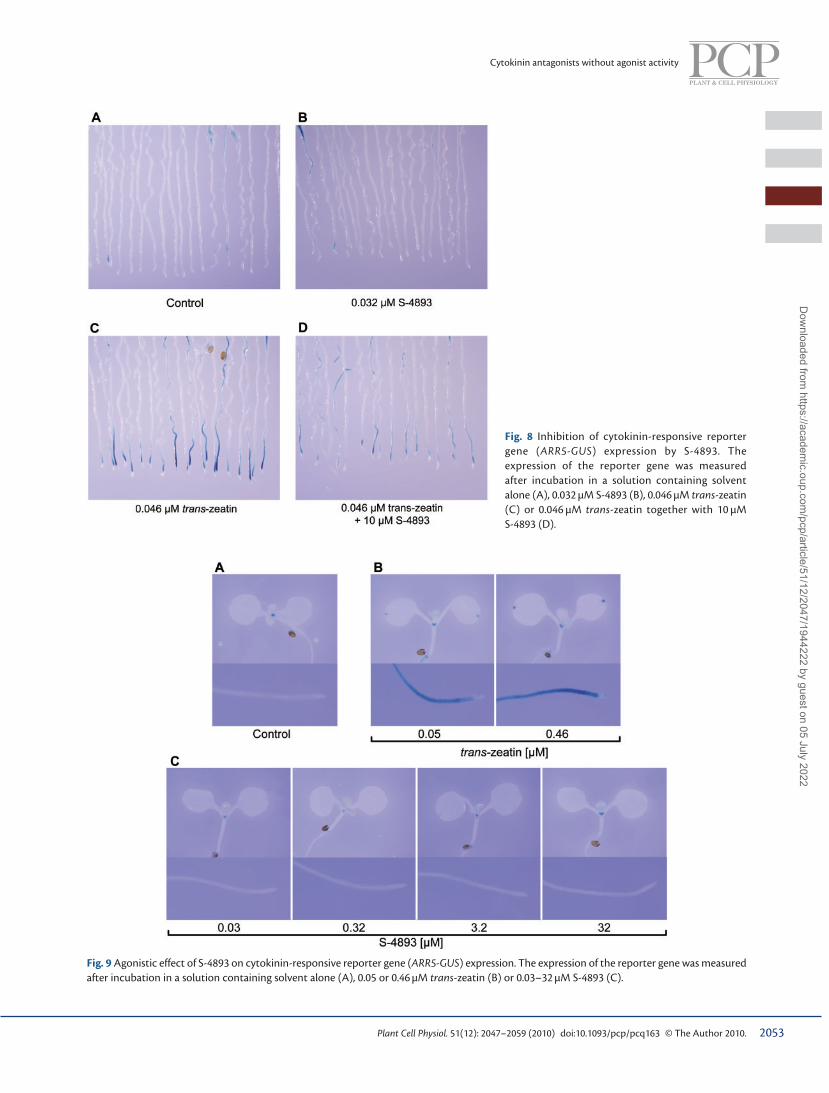
Fig. 8 Inhibition of cytokinin-responsive reporter gene ( ARR5-GUS ) expression by S-4893. The expression of the reporter gene was measured after incubation in a solution containing solvent alone (A), 0.032 µM S-4893 (B), 0.046 µM trans -zeatin (C) or 0.046 µM trans -zeatin together with 10 µM S-4893 (D).
Fig. 9 Agonistic effect of S-4893 on cytokinin-responsive reporter gene ( ARR5-GUS ) expression. The expression of the reporter gene was measured after incubation in a solution containing solvent alone (A), 0.05 or 0.46 µM trans -zeatin (B) or 0.03–32 µM S-4893 (C).
2053
Cytokinin antagonists without agonist activity
Plant Cell Physiol. 51(12): 2047–2059 (2010) doi:10.1093/pcp/pcq163 © The Author 2010.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/51/12/2047/1944222 by guest on 05 July 2022
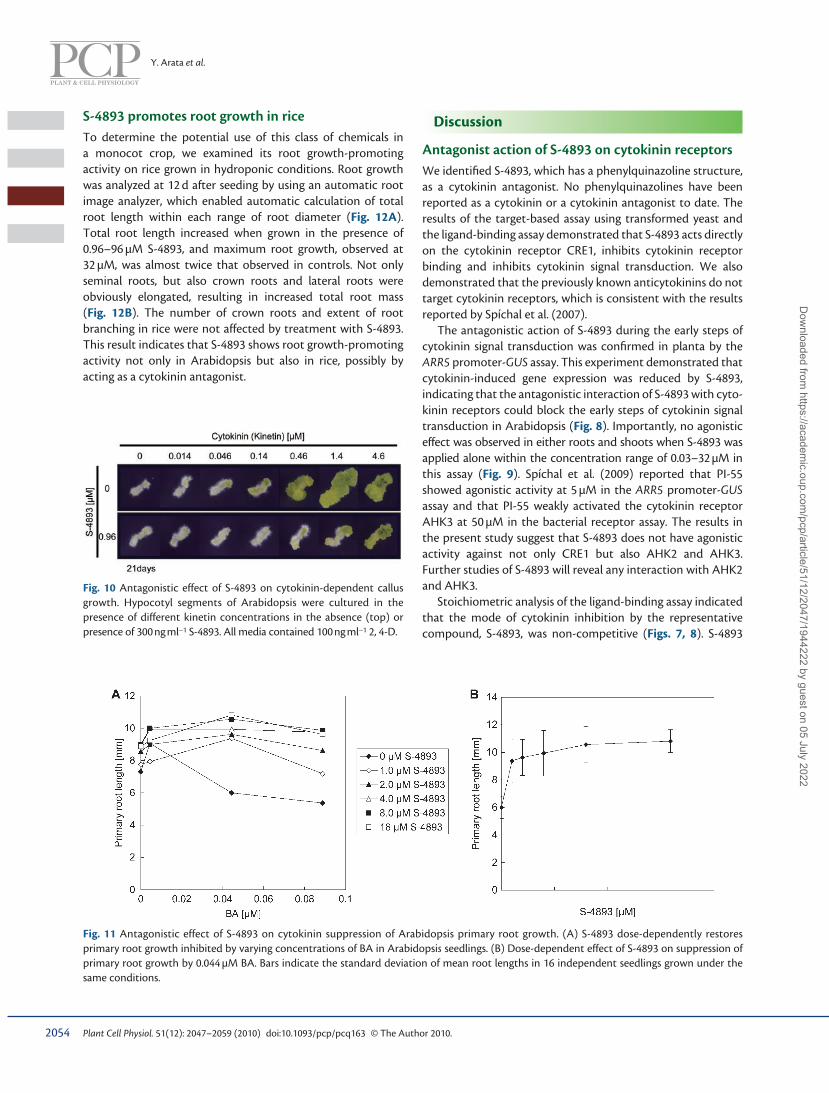
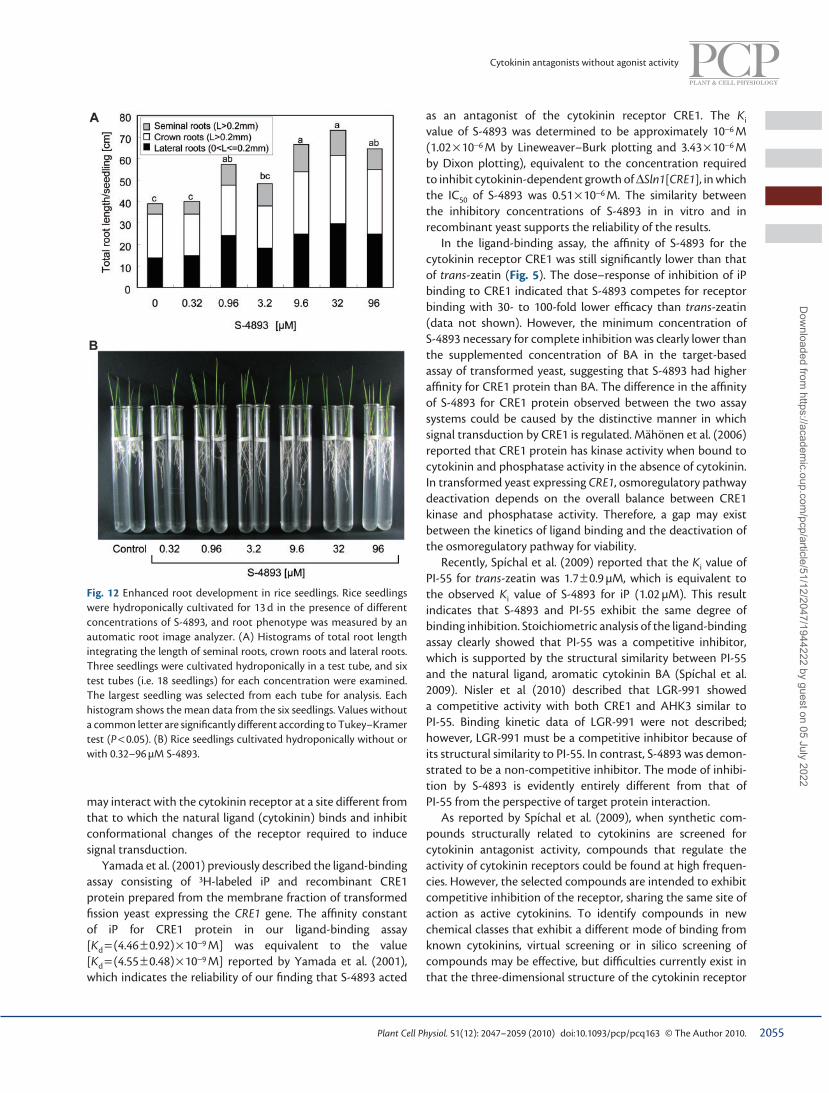
S-4893 promotes root growth in rice To determine the potential use of this class of chemicals in a monocot crop, we examined its root growth-promoting activity on rice grown in hydroponic conditions. Root growth was analyzed at 12 d after seeding by using an automatic root image analyzer, which enabled automatic calculation of total root length within each range of root diameter ( Fig. 12A ). Total root length increased when grown in the presence of 0.96–96 µM S-4893, and maximum root growth, observed at 32 µM, was almost twice that observed in controls. Not only seminal roots, but also crown roots and lateral roots were obviously elongated, resulting in increased total root mass ( Fig. 12B ). The number of crown roots and extent of root branching in rice were not affected by treatment with S-4893. This result indicates that S-4893 shows root growth-promoting activity not only in Arabidopsis but also in rice, possibly by acting as a cytokinin antagonist.
Fig. 10 Antagonistic effect of S-4893 on cytokinin-dependent callus growth. Hypocotyl segments of Arabidopsis were cultured in the presence of different kinetin concentrations in the absence (top) or presence of 300 ng ml − 1 S-4893. All media contained 100 ng ml − 1 2, 4-D.
Fig. 11 Antagonistic effect of S-4893 on cytokinin suppression of Arabidopsis primary root growth. (A) S-4893 dose-dependently restores primary root growth inhibited by varying concentrations of BA in Arabidopsi s seedlings. (B) Dose-dependent effect of S-4893 on suppression of primary root growth by 0.044 µM BA. Bars indicate the standard deviation of mean root lengths in 16 independent seedlings grown under the same conditions.
Discussion
Antagonist action of S-4893 on cytokinin receptors We identifi ed S-4893, which has a phenylquinazoline structure, as a cytokinin antagonist. No phenylquinazolines have been reported as a cytokinin or a cytokinin antagonist to date. The results of the target-based assay using transformed yeast and the ligand-binding assay demonstrated that S-4893 acts directly on the cytokinin receptor CRE1, inhibits cytokinin receptor binding and inhibits cytokinin signal transduction. We also demonstrated that the previously known anticytokinins do not target cytokinin receptors, which is consistent with the results reported by Spíchal et al. (2007) .
The antagonistic action of S-4893 during the early steps of cytokinin signal transduction was confi rmed in planta by the ARR5 promoter- GUS assay. This experiment demonstrated that cytokinin-induced gene expression was reduced by S-4893, indicating that the antagonistic interaction of S-4893 with cyto-kinin receptors could block the early steps of cytokinin signal transduction in Arabidopsis ( Fig. 8 ). Importantly, no agonistic effect was observed in either roots and shoots when S-4893 was applied alone within the concentration range of 0.03–32 µM in this assay ( Fig. 9 ). Spíchal et al. (2009) reported that PI-55 showed agonistic activity at 5 µM in the ARR5 promoter- GUS assay and that PI-55 weakly activated the cytokinin receptor AHK3 at 50 µM in the bacterial receptor assay. The results in the present study suggest that S-4893 does not have agonistic activity against not only CRE1 but also AHK2 and AHK3. Further studies of S-4893 will reveal any interaction with AHK2 and AHK3.
Stoichiometric analysis of the ligand-binding assay indicated that the mode of cytokinin inhibition by the representative compound, S-4893, was non-competitive ( Figs. 7 , 8 ). S-4893
2054
Y. Arata et al.
Plant Cell Physiol. 51(12): 2047–2059 (2010) doi:10.1093/pcp/pcq163 © The Author 2010.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/51/12/2047/1944222 by guest on 05 July 2022
may interact with the cytokinin receptor at a site different from that to which the natural ligand (cytokinin) binds and inhibit conformational changes of the receptor required to induce signal transduction.
Yamada et al. (2001) previously described the ligand-binding assay consisting of 3 H-labeled iP and recombinant CRE1 protein prepared from the membrane fraction of transformed fi ssion yeast expressing the CRE1 gene. The affi nity constant of iP for CRE1 protein in our ligand-binding assay [ K d = (4.46 ± 0.92) × 10 − 9 M] was equivalent to the value [ K d = (4.55 ± 0.48) × 10 − 9 M] reported by Yamada et al. (2001) , which indicates the reliability of our fi nding that S-4893 acted
Fig. 12 Enhanced root development in rice seedlings. Rice seedlings were hydroponically cultivated for 13 d in the presence of different concentrations of S-4893, and root phenotype was measured by an automatic root image analyzer. (A) Histograms of total root length integrating the length of seminal roots, crown roots and lateral roots. Three seedlings were cultivated hydroponically in a test tube, and six test tubes (i.e. 18 seedlings) for each concentration were examined. The largest seedling was selected from each tube for analysis. Each histogram shows the mean data from the six seedlings. Values without a common letter are signifi cantly different according to Tukey–Kramer test ( P < 0.05). (B) Rice seedlings cultivated hydroponically without or with 0.32–96 µM S-4893.
as an antagonist of the cytokinin receptor CRE1. The K i value of S-4893 was determined to be approximately 10 − 6 M (1.02 × 10 − 6 M by Lineweaver–Burk plotting and 3.43 × 10 − 6 M by Dixon plotting), equivalent to the concentration required to inhibit cytokinin-dependent growth of ∆ Sln1 [ CRE1 ], in which the IC 50 of S-4893 was 0.51 × 10 − 6 M. The similarity between the inhibitory concentrations of S-4893 in in vitro and in recombinant yeast supports the reliability of the results.
In the ligand-binding assay, the affi nity of S-4893 for the cytokinin receptor CRE1 was still signifi cantly lower than that of trans -zeatin ( Fig. 5 ). The dose–response of inhibition of iP binding to CRE1 indicated that S-4893 competes for receptor binding with 30- to 100-fold lower effi cacy than trans -zeatin (data not shown). However, the minimum concentration of S-4893 necessary for complete inhibition was clearly lower than the supplemented concentration of BA in the target-based assay of transformed yeast, suggesting that S-4893 had higher affi nity for CRE1 protein than BA. The difference in the affi nity of S-4893 for CRE1 protein observed between the two assay systems could be caused by the distinctive manner in which signal transduction by CRE1 is regulated. Mähönen et al. (2006) reported that CRE1 protein has kinase activity when bound to cytokinin and phosphatase activity in the absence of cytokinin. In transformed yeast expressing CRE1 , osmoregulatory pathway deactivation depends on the overall balance between CRE1 kinase and phosphatase activity. Therefore, a gap may exist between the kinetics of ligand binding and the deactivation of the osmoregulatory pathway for viability.
Recently, Spíchal et al. (2009) reported that the K i value of PI-55 for trans -zeatin was 1.7 ± 0.9 µM, which is equivalent to the observed K i value of S-4893 for iP (1.02 µM). This result indicates that S-4893 and PI-55 exhibit the same degree of binding inhibition. Stoichiometric analysis of the ligand-binding assay clearly showed that PI-55 was a competitive inhibitor, which is supported by the structural similarity between PI-55 and the natural ligand, aromatic cytokinin BA ( Spíchal et al. 2009 ). Nisler et al (2010) described that LGR-991 showed a competitive activity with both CRE1 and AHK3 similar to PI-55. Binding kinetic data of LGR-991 were not described; however, LGR-991 must be a competitive inhibitor because of its structural similarity to PI-55. In contrast, S-4893 was demon-strated to be a non-competitive inhibitor. The mode of inhibi-tion by S-4893 is evidently entirely different from that of PI-55 from the perspective of target protein interaction.
As reported by Spíchal et al. (2009) , when synthetic com-pounds structurally related to cytokinins are screened for cytokinin antagonist activity, compounds that regulate the activity of cytokinin receptors could be found at high frequen-cies. However, the selected compounds are intended to exhibit competitive inhibition of the receptor, sharing the same site of action as active cytokinins. To identify compounds in new chemical classes that exhibit a different mode of binding from known cytokinins, virtual screening or in silico screening of compounds may be effective, but diffi culties currently exist in that the three-dimensional structure of the cytokinin receptor
2055
Cytokinin antagonists without agonist activity
Plant Cell Physiol. 51(12): 2047–2059 (2010) doi:10.1093/pcp/pcq163 © The Author 2010.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/51/12/2047/1944222 by guest on 05 July 2022

protein at the ligand-binding site has not yet been determined. Therefore, as shown by our study, screening of a large number of chemicals by target-based assay is an effective method to identify chemicals that exhibit a different mode of binding from cytokinins, although the effi ciency of the method is rela-tively low. Only two compounds that inhibited cytokinin action in the target-based assay of the yeast system were selected from approximately 80,000 compounds in our chemical library. This extremely low hit rate in the yeast system may be due to the strict structural requirements of ligand binding to cytokinin receptors. As our chemical library used for the screening of cytokinin antagonists has high structural diversity, it is reason-able that a limited number of active compounds were identi-fi ed in our chemical library.
Physiological responses of plants to S-4893 Cytokinins are known to affect root growth negatively, and cytokinin antagonists are therefore expected to promote root growth. In the present study, we screened for compounds that exhibit antagonistic activity toward CRE1, which mainly functions in root tissues, and, as expected, these compounds showed root growth-promoting activity. S-4893, which showed enhanced antagonistic activity toward CRE1, also showed enhanced root growth-promoting activity. This result also suggests that antagonistic to CRE1 activity is correlated with the effect on root growth promotion. These results clearly show that antagonistic action on cytokinin signaling is one potential MOA to induce root growth promotion. S-4893 inhibited cyto-kinin effects not only on root elongation but also on callus for-mation, in agreement with its targeting of cytokinin receptors.
We also demonstrated that root growth promotion by S-4893 was achieved not only in Arabidopsis but also in rice ( Fig. 12 ). S-4893 promoted seminal, crown and lateral root growth in rice under hydroponic culture conditions, possibly due to the antagonistic action of S-4893 on cytokinin receptors in rice. This fi nding is supported by the identifi cation of cytokinin receptors that show high homology with those of Arabidopsis in rice ( Du et al. 2007 ); a similar cytokinin-sensing mechanism to that mediated by Arabidopsis CRE1 may be active in rice. This result further suggests that S-4893 could potentially promote root growth in various agronomically important plants. Spíchal et al. (2009) and Nisler et al. (2010) also reported that PI-55 and LGR-991, respectively, antagonized cytokinin effects in bioassays with various plant species. These results suggest that cytokinin antagonists might be widely applicable as PGRs to modify the traits of a range of plants.
Spíchal et al. (2009) reported that PI-55, which has a pre-dominantly antagonistic activity on CRE1, increased primary root length in AHK2/AHK3 double mutants but not in wild-type plants. PI-55 also exhibited antagonistic activity against the effects of exogenous and endogenous cytokinins on root branching in both wild-type and AHK2/AHK3 double-mutant Arabidopsis. In contrast, S-4893 increased primary root length ( Table 2 , Fig. 11 ), but did not have a positive effect on root branching in wild-type Arabidopsis (data not shown).
Three cytokinin receptors have been reported to regulate primary root elongation and root branching ( Riefl er et al. 2006 ). The antagonistic activity of S-4893 on AHK2 and AHK3 may differ from that of PI-55. The difference in root growth pheno-type observed in both studies may also be due to differences in culture conditions or other methodological factors.
Because S-4893 inhibits cytokinin responses by targeting cytokinin receptors without exhibiting any recognizable ago-nist activity, it will be a useful tool in the study of plant physiol-ogy. It also has potential for practical applications in agriculture to improve the root systems of various crops, although further studies of S-4893 and derivatives regarding interaction with other cytokinin receptors in various crops, in-depth toxicity testing and in-depth environmental testing will have to be carried out to put them to practical use.
Materials and Methods
Target-based assay for a cytokinin receptor The chemical library used as a screening source was obtained from commercial sources and consisted of aproximately 80,000 chemicals for target-based assay of a yeast system. The trans-formed yeast strain TM182 ( Maeda et al. 1994 ) carrying the vector p415CYC-CRE1 ( Inoue et al. 2001 ), which consisted of the A. thaliana CRE1 gene under the control of a yeast CYC pro-moter, was used in a target-based assay. A wild strain of yeast, AH22 ( Gunge and Sakaguchi 1981 ), was used as a control. The transformed yeast was pre-cultured overnight in a Gal-Ura-Leu medium that contained 6.7 g l − 1 yeast nitrogen base without amino acids (Difco), 1.66 g l − 1 SC-HIS-LEU-URA (Q-BIOgene), 0.076 g l − 1 histidine and 20 g l − 1 galactose. YPD medium that included 10 g l − 1 yeast extract, 20 g l − 1 bacto-peptone and 20 g l − 1 glucose was used for pre-culture of wild yeast. For cul-ture in the target-based assay, the pre-cultured suspension of the transformed yeast was inoculated at 0.5 % into fresh Glu-Ura-Leu medium that contained 6.7 g l − 1 yeast nitrogen base without amino acids (Difco), 1.66 g l − 1 SC-HIS-LEU-URA (Q-BIOgene), 0.076 g l − 1 histidine and 20 g l − 1 glucose. The assay for antagonists was conducted in 96-well microplates, with each well containing a 100 µl aliquot of the inoculated medium, 8.9 µM (2 ppm) BA and the indicated concentration of the test compound. The assay for agonists was conducted like the assay for antagonists but without BA. The transformed yeast cells were cultivated at 30 ° C in 96-well microplates without any agitation for 1 d. The optical density of each well was measured at 600 nm by an automatic microplate reader (Corona electric MTP-300). The control assay was also carried out with the wild strain in YPD medium under conditions similar to those used for the transformed yeast. Relative growth was determined as the percentage of yeast growth for the test compound compared with that for solvent control [1 % dimethylsulfoxide (DMSO)]. The IC 50 of the test compound was calculated from a dose–response curve by XLfi t software (ID Business Solutions Ltd.).
2056
Y. Arata et al.
Plant Cell Physiol. 51(12): 2047–2059 (2010) doi:10.1093/pcp/pcq163 © The Author 2010.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/51/12/2047/1944222 by guest on 05 July 2022

Bioassay for screening for root growth promoters The chemical library used as a screening source was obtained from commercial sources and consisted of 37,921 chemicals for bioassay for RGPs. All test compounds were prepared as a solution of DMSO at 1,500 ppm. The bioassay for RGPs was constructed in a cluster tube with a diameter of 9.0 mm and height of 44.5 mm (BM4140, BIO MEDICAL EQUIPMENT) con-taining 600 µl of solid medium supplemented with the test compound at a fi nal concentration of 2 or 10 ppm per tube and eight tubes per treatment. The solid medium (ENSHI solid medium) contained 950 mg l − 1 calcium nitrate, 810 mg l − 1 potas-sium nitrate, 500 mg l − 1 magnesium sulfate, 155 mg l − 1 ammo-nium dihydrogen phosphate, 22.6 mg l − 1 Fe-EDTA, 2.86 mg l − 1 boric acid, 1.82 mg l − 1 manganese(II) sulfate, 0.22 mg l − 1 zinc sulfate, 0.08 mg l − 1 copper(II) sulfate, 0.025 mg l − 1 sodium molybdate and 8 g l − 1 agar. The pH of the medium was adjusted to 6.0 with 1 N KOH. Approximately 25–50 sterilized seeds of A. thaliana ecotype Columbia per tube were planted on the solid medium and cultivated at 22 ° C under periodic illumina-tion with 16 h light/8 h dark intervals. The primary root length was measured at 10 d after seeding. The compounds were pri-marily selected based on root growth promotion > 120 % of the control. Root growth was calculated as the mean value of the eight respective mean lengths of primary roots in eight respective tubes.
Preparation of S-4893 and anticytokinins S-4893 was chemically synthesized from 2,6-dichloro-4-phenylquinazoline and 3-amino-1-propanol according to the procedure reported by Kwon et al. (1973) . AntiCK-1, 4-benzylamino-2-methylthiopyrido[2,3- d ]pyrimidine, was obtained through a custom synthesis by a commercial service provider (Sumika Technoservice Corporation). The compound was prepared from 4-oxo-2-thionodihydro-2 H -pyrido[2,3- d ]pyrimidine ( Stanovnik and Tisler 1972 ) essentially according to the chemical process described by Koren et al. (1987) . The following compounds were purchased from commercial sources: AntiCK-2, 3-benzyl-7-hydroxy-5-methyl-3 H -1,2,3-triazolo[4,5- d ]pyrimidine (ASINEX); AntiCK-3, 1-benzyl-3-(4-chlorophenyl)urea (SALOR); AntiCK-4, propyl N -(3,4-dichlorophenyl)carbamate (Sigma); and AntiCK-5, ethyl 5-amino-1-benzylpyrazole-4-carboxylate (ASINEX).
Hydroponic culture of rice seedlings Rice seeds ( Oryza sativa cv Nipponbare) were immersed in water at 30 ° C for 2 d to stimulate germination. Three treated seeds were placed on a fl oat and cultivated hydroponically in a test tube (25 mm × 200 mm) containing 50 ml of Kimura B nutrient solution at 1/8th strength ( Yoshida et al. 1972 ) supple-mented or not with 0.32–96 µM S-4893. Light exposure of the root zones was prevented by wrapping the tubes with alumi-num foil, and the rice seedlings were grown at 25 ° C under continuous light for 13 d. The root phenotype was analyzed by an automatic root image analyzer (WinRHIZO, Regent Instruments Inc.). The seminal and crown roots corresponded
to roots > 0.2 mm in diameter, and lateral roots corresponded to roots < 0.2 mm in diameter. The length of seminal roots was also measured, and the length of crown roots was calculated by subtracting the seminal root length from total root length > 0.2 mm in diameter. The reproducibility of the root pheno-type was confi rmed by duplication of the experiment.
In vitro ligand-binding assay To prepare CRE1 recombinant protein for the in vitro binding assay, the high expression plasmid p425GPD-CRE1 was con-structed by cloning the CRE1 gene of A. thaliana into the Bam HI site of the high copy vector p425GPD ( Mumberg et al. 1995 ). Using the plasmid p415CYC-CRE1 as template DNA, full-length CRE1 cDNA was amplifi ed by PCR with 5 ′ and 3 ′ oligonucleotide primers, under the conditions described in the manufacturer's instructions for thermostable KOD-Plus DNA polymerase (Toyobo). The nucleotide sequences of the sense and antisense primers were 5 ′ -CGGGATCCATGAACTGGGCACTCAACAATCATC-3 ′ and 5 ′ -GCTCTAGATTACGACGAAGGTGAGATAGGATTAG-3 ′ , respectively. The cell mass of the yeast strain TM182 transformed with p425GPD-CRE1 was harvested for the pro-tein fraction containing recombinant CRE1 protein, which was essential for the in vitro binding assay. A cell mass of approximately 3.2 ml, obtained from 400 ml of cultured suspen-sion of the transformed yeast, was repeatedly washed and sus-pended with 8.0 ml of cold phosphate buffer (50 mM) and fi nally resuspended in 9.6 ml of the same buffer supplemented with 5 mM dithiothreitol and 0.5 mM phenylmethylsulfonyl fl uoride. Yeast cells were disrupted by shaking a microtube containing a 200 µl aliquot of the suspension with glass beads (diameter, 0.25–0.5 mm) for three 30 s intervals in a Multi-beads shocker MB-200 (Yasui Kikai). Glass beads were separated from the suspension of disrupted yeast cells by centrifugation at 1,500 × g for 10 min. A pellet of the transformed yeast cells was collected by re-centrifugation of the supernatant at 100,000 × g for 1 h. The membrane fraction was prepared as a suspension of the pellet in cold 50 mM phosphate buffer containing 1 % sucrose monocaprate. The protein content in the membrane fraction was determined by Bradford protein assay with bovine serum albumin as a standard. For the in vitro binding assay, 150 µg of the membrane fraction containing recombinant CRE1 protein, the indicated concentration of 3 H-labeled iP and a test compound were mixed and incubated on ice for 1 h. After incubation for ligand binding, the membrane fraction was harvested and washed in cold 50 mM phosphate buffer on a Whatmann GF/B glass fi ber fi lter in a cell harvester (Brandel M-24). Ligand binding was measured as the radioactivity of the membrane fraction on the fi lter determined by a liquid scintillation counter (PerkinElmer Tri-Carb 2900TR, Inc.). An ethanol solution of 3 H-labeled iP (37.0 MBq ml − 1 ) was pur-chased from Amersham Biosciences. The specifi c radioactivity of 3 H-labeled iP was 74.0 GBq mmol − 1 . Kinetic analysis of the obtained data was conducted according to Lineweaver–Burk plotting ( Lineweaver and Burk 1934 ) and Dixon plotting ( Dixon and Webb 1979 ).
2057
Cytokinin antagonists without agonist activity
Plant Cell Physiol. 51(12): 2047–2059 (2010) doi:10.1093/pcp/pcq163 © The Author 2010.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/51/12/2047/1944222 by guest on 05 July 2022

In vivo assay for callus induction in Arabidopsis Sterilized seeds of A. thaliana ecotype Landsberg erecta were planted on a solid germination medium containing Murashige and Skoog salt mixture (Wako Pure Chemical Industries, Ltd.), 0.05 % MES-KOH (pH 5.7), 1 % sucrose, 100 mg l − 1 inositol, 10 mg l − 1 thiamine-HCl, 1 mg l − 1 nicotinic acid, 1 mg l − 1 pyridox-ine-HCl and 0.3 % Phytagel (Sigma-Aldrich). The seedlings were obtained after cultivation at 22 ° C for 8 d under constant illumination. Callus formation from excised hypocotyls of the seedlings was induced by cultivation on germination medium supplemented with 2,4-D (100 µg l − 1 ) and kinetin (the indicated concentration). The effect of addition of S-4893 to the medium on the callus induction phenotype was observed after cultivation at 22 ° C for 21 d.
In vitro assay for root elongation in Arabidopsis Sterilized seeds of A. thaliana ecotype Columbia were planted on ENSHI solid medium supplemented with the indicated con-centrations of BA and S-4893 in 1.4 ml cluster tubes (Matrix). Seedlings were grown on ENSHI solid medium under continu-ous illumination at 22 ° C, and primary root length was mea-sured at 10 d after seeding. Root growth was calculated as the mean length of primary roots of 16 tubes, and the reproduc-ibility of the results was confi rmed by duplication of the assay.
Reporter gene assay in transgenic Arabidopsis Transgenic Arabidopsis with a chimeric gene comprising the cytokinin-inducible promoter of the ARR5 gene and a bacterial GUS reporter gene was prepared basically according to the method reported by D'Agostino et al. (2000) . A region upstream of ARR5 was PCR amplifi ed with the primers 5 ′ -CCCAAGCTTGGAAACCAATAAAGCATATTTG-3 ′ and 5 ′ -TCCCCCGGGCTGAGAGATGAGAGGAGAATAA-3 ′ , digested with Hin dIII and Sma I and then cloned between the Hin dIII and Sma I sites of pGPTV-Bar ( Becker et al. 1992 ). Control transgenic Arabidopsis with the 35S promoter gene instead of the ARR5 promoter gene was also prepared by a similar procedure to that reported by D'Agostino et al. (2000) . Sterilized seeds of the transgenic Arabidopsis were planted on solid MS medium and cultivated at 22 ° C under continuous illumination for 7 d. Histochemical analysis of the GUS marker enzyme was performed essentially according to Jefferson et al. (1987) as modifi ed by Iida et al. (1990) . The seedlings were incubated in sterilized water supple-mented with the indicated concentration of trans -zeatin and S-4893 at 22 ° C under continuous illumination for 1 h in the competitive assay. The seedlings were washed and reincubated in fresh sterilized water under the same conditions for 3 h. In the assay to examine the cytokinin agonist activity of S-4893, the seedlings were incubated overnight in sterilized water sup-plemented with the indicated concentration of trans -zeatin or S-4893. After replacing the sterilized water with a staining solu-tion containing 5-bromo-4-chloro-3-indolyl- β - D -glucuronic acid, the seedlings were incubated overnight at 37 ° C in the dark. Endogenous pigments were destained with 70 % ethanol, and GUS staining in the root and shoot tissue of seedlings
was recorded with a cooled CCD camera (Keyence VB-7010) attached to a fl uorescence stereoscopic microscope (Keyence VB-G05). The reporter gene assay was duplicated to confi rm the reproducibility of the results.
Acknowledgments
The authors are grateful to Dr. Higuchi for providing some materials described in this study and valuable comments. We also thank our colleagues and technical staff for their many years of effort.
References
Becker , D. , Kemper , E. , Schell , J. and Masterson , R. ( 1992 ) New plant binary vectors with selectable markers located proximal to the left T-DNA border . Plant Mol. Biol. 20 : 1195 – 1197 .
Benková , E. and Hejátko , J. ( 2009 ) Hormone interactions at the root apical meristem . Plant Mol. Biol. 69 : 383 – 396 .
Choi , J. and Hwang , I. ( 2007 ) Cytokinin: perception, signal transduction, and role in plant growth and development . J. Plant Biol. 50 : 98 – 108 .
D'Agostino , I.B. , Deruère , J. and Kieber , J.J. ( 2000 ) Characterization of the response of the Arabidopsis response regulator gene family to cytokinin . Plant Physiol. 124 : 1706 – 1717 .
Dello Ioio , R. , Linhares , F.S. , Scacchi , E. , Casamitjana-Martinez , E. , Heidstra , R. , Costantino , P. , et al . ( 2007 ) Cytokinins determine Arabidopsis root-meristem size by controlling cell differentiation . Curr. Biol. 17 : 678 – 682 .
Ding , Y. and Ma , Q.-H. ( 2006 ) Biosynthesis, signal transduction and molecular regulation of cytokinins . Curr. Pharm. Anal. 2 : 169 – 182 .
Dixon , M. and Webb , E.C. ( 1979 ) Enzymes . Longman , London . Du , L. , Jiao , F. , Chu , J. , Gulei J. , Chen , M. and Wu , P. ( 2007 ) The two-
component signal system in rice (Oryza sativa L.): a genome-wide study of cytokinin signal perception and transduction . Genomics 89 : 697 – 707 .
Ferguson , L. and Lessenger J.E. ( 2006 ) Plant growth regulators . In Agricultural Medicine . Edited by Lessenger J.E. pp. 156 – 166 . Springer , New York .
Ferreira , F.J. and Kieber , J.J. ( 2005 ) Cytokinin signaling . Curr. Opin. Plant
Biol. 8 : 518 – 525 . Gunge , N. and Sakaguchi , K. ( 1981 ) Intergeneric transfer of
deoxyribonucleic acid killer plasmids, pGKl1 and pGKl2, from Kluyveromyces lactis into Saccharomyces cerevisiae by cell fusion . J. Bacteriol. 147 : 155 – 160 .
Hecht , S.M. , Bock , R.M. , Schmitz , R.Y. , Skoog , F. and Leonard , N.J. ( 1971 ) Cytokinins: development of a potent antagonist . Proc. Natl Acad.
Sci. USA 68 : 2608 – 2610 . Higuchi , M. , Pischke , M.S. , Mahoen , A.P. , Miyawaki , K. , Hashimoto , Y. ,
Seki , M. , et al . ( 2004 ) In planta functions of the Arabidopsis cytokinin receptor family . Proc. Natl Acad. Sci. USA 101 : 8821 – 8826 .
Hwang , I. and Sakakibara , H. ( 2006 ) Cytokinin biosynthesis and perception . Physiol. Plant. 126 : 528 – 583 .
Iida , A. , Seki , M. , Kamada , M. , Yamada , Y. and Morikawa , H. ( 1990 ) Gene delivery into cultured plant cells by DNA-coated gold particles accelerated by a pneumatic particle gun . Theor. Appl. Genet. 80 : 813 – 816 .
2058
Y. Arata et al.
Plant Cell Physiol. 51(12): 2047–2059 (2010) doi:10.1093/pcp/pcq163 © The Author 2010.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/51/12/2047/1944222 by guest on 05 July 2022

Inoue , T. , Higuchi , M. , Hashimoto , Y. , Seki , M. , Kobayashi , M. , Kato , T. , et al . ( 2001 ) Identifi cation of CRE1 as a cytokinin receptor from Arabidopsis . Nature 409 : 1060 – 1063 .
Iwamura , H. ( 1994 ) Cytokinin antagonists: synthesis and biological activity . In Cytokinins: Chemistry, Activity and Function . Edited by Mok , D.W.S. and Mok , M.C. pp. 43 – 55 . CRC Press , Boca Raton, FL .
Iwamura , H. , Masuda , N. , Koshimizu , K. and Matsubara , S. ( 1983 ) Quantitative aspects of the receptor binding of cytokinin agonists and antagonists . J. Med. Chem. 26 : 838 – 844 .
Iwamura , H. , Murakami , S. , Koga , J. , Matsubarab , S. and Koshimizu , K. ( 1979 ) Quantitative analysis of anticytokinin activity of 4-substituted 2-methylthiopyrido[2,3-d]pyrimidines . Phytochemistry 18 : 1265 – 1268 .
Iwamura , H. , Murakami , S. , Koshimizu , K. and Matsubara , S. ( 1985 ) Quantitative structure–activity relationships in cytokinin agonistic and antagonistic pyrido[2,3-d]pyrimidine derivatives: insights into receptor topology . J. Med. Chem. 28 : 577 – 583 .
Jefferson , R.A. , Kavanagh , T.A. and Bevan , M.W. ( 1987 ) GUS fusions: β -glucuronidase as a sensitive and versatile gene fusion marker in higher plants . EMBO J. 6 : 3901 – 3907 .
Kakimoto , T. ( 2001 ) Identifi cation of plant cytokinin biosynthetic enzymes as dimethylallyl diphosphate:ATP/ADP isopentenyltrans-ferases . Plant Cell Physiol. 42 : 677 – 685 .
Kakimoto , T. ( 2003 ) Perception and signal transduction of cytokinins . Annu. Rev. Plant Biol. 54 : 605 – 627 .
Kakimoto , T. and Higuchi , M. ( 2005 ) Transformed cell co-expressing cytokinin receptor and cytokinin biosynthesis enzyme. European Patent Application EP1522586 .
Kakimoto , T. , Higuchi , M. and Inoue , T. ( 2002 ) Analysis of agonist-activity and antagonist-activity to cytokinin receptor. European Patent Application EP1241182 .
Karanov , E. , Alexieva , V. , Golovinsky , E. and Haimova , M. ( 1993 ) Cytokinin and anticytokinin activity of some 4-substituted 1H-pyrazoles and 8-aza analogues of adenine . Plant Growth Regul. 13 : 7 – 11 .
Kefford , N.P. , Zwar , J.A. and Bruce , M.I. ( 1968 ) Antagonism of purine and urea cytokinin activities by derivatives of benzylurea . In Biochemistry and Physiology of Plant Growth Substances . Edited by Wightman , F. and Setterfi eld , G. pp. 61 – 69 . Runge Press Ltd. , Ottawa .
Koren , B. , Stanovnik , B. and Tisler , M. ( 1987 ) Transformations of 1,2,4-thiadiazolo[2,3-x]azines. Heterocycles 26 : 689 – 697 .
Koshioka , M. , Takeuchi , Y. , Saka , H. and Kamuro , Y. ( 2005 ) Mutual and close relationship between basic studies and practical application on plant growth substances . Regul. Plant Growth Dev. 40 : 111 – 123 .
Kuroha , T. , Ueguchi , C. , Sakakibara , H. and Satoh , S. ( 2006 ) Cytokinin receptors are required for normal development of auxin-transporting vascular tissues in the hypocotyl but not in adventitious roots . Plant Cell Physiol. 47 : 234 – 243 .
Kwon , S. , Ikeda , F. and Isagawa , K. ( 1973 ) Cyclization of quinazoline derivatives . Nippon Kagaku Kaishi 10 : 1944 – 1948 .
Kyozuka , J. ( 2007 ) Control of shoot and root meristem function by cytokinin . Curr. Opin. Plant Biol. 10 : 442 – 446 .
Li , X. , Mo , X. , Shou , H. and Wu , P. ( 2006 ) Cytokinin-mediated cell cycling arrest of pericycle founder cells in lateral root initiation of Arabidopsis . Plant Cell Physiol. 47 : 1112 – 1123 .
Lineweaver , H. and Burk , D. ( 1934 ) The determination of enzyme dissociation constants . J. Amer. Chem. Soc. 56 : 658 – 666 .
Maeda , T. , Wurgler-Murphy , S.M. and Saito , H. ( 1994 ) A two-component system that regulates an osmosensing MAP kinase cascade in yeast . Nature 369 : 187 – 188 .
Mähönen , A.P. , Higuchi , M. , Törmäkangas , K. , Miyawaki , K. , Pischke , M.S. , Sussman , M.R. , et al . ( 2006 ) Cytokinins regulate a bidirectional phosphorelay network in Arabidopsis . Curr. Biol. 11 : 1116 – 1122 .
Miyawaki , K. , Tarkowski , P. , Matsumoto-Kitano , M. , Kato , T. , Sato , S. , Tarkowska , D. , et al . ( 2006 ) Roles of Arabidopsis ATP/ADP isopentenyltransferases and tRNA isopentenyltransferases in cytokinin biosynthesis . Proc. Natl Acad. Sci. USA 103 : 16598 – 16603 .
Mumberg , D. , Müller , R. and Funk , M. ( 1995 ) Yeast vectors for the controlled expression of heterologous proteins in different genetic backgrounds . Gene 156 : 119 – 122 .
Nishimura , C. , Ohashi , Y. , Sato , S. , Kato , T. , Tabata , S. and Ueguchi , C. ( 2004 ) Histidine kinase homologs that act as cytokinin receptors possess overlapping functions in the regulation of shoot and root growth in Arabidopsis . Plant Cell 16 : 1365 – 1377 .
Nisler , J. , Zatloukal , M. , Popa , I. , Doležal , K. , Strnad , M. and Spíchal , L. ( 2010 ) Cytokinin receptor antagonists derived from 6-benzylaminopurine . Phytochemistry 71 : 823 – 830 .
Riefl er , M. , Novak , O. , Strnad , M. and Schmülling , T. ( 2006 ) Arabidopsis cytokinin receptor mutants reveal functions in shoot growth, leaf senescence, seed size, germination, root development, and cytokinin metabolism . Plant Cell 18 : 40 – 54 .
Romanov , G.A. , Lomin , S.N. and Schmülling , T. ( 2006 ) Biochemical characteristics and ligand-binding properties of Arabidopsis cytokinin receptor AHK3 compared to CRE1/AHK4 as revealed by a direct binding assay . J. Exp. Bot. 57 : 4051 – 4040 .
Romanov , G.A. , Spíchal , L. , Lomin , S.N. , Strnad , M. and Schmülling , T. ( 2005 ) A live cell hormone-binding assay on transgenic bacteria expressing a eukaryotic receptor protein . Anal. Biochem. 347 : 129 – 134 .
Spíchal , L. , Krystof , V. , Paprskárová , M. , Lenobel , R. , Stýskala , J. , Binarová , P. , et al . ( 2007 ) Classical anticytokinins do not interact with cytokinin receptors but inhibit cyclin-dependent kinases . J. Biol. Chem. 282 : 14356 – 14363 .
Spíchal , L. , Werner , T. , Popa , I. , Riefl er , M. , Schmülling , T. and Strnad , M. ( 2009 ) The purine derivative PI-55 blocks cytokinin action via receptor inhibition . FEBS J. 276 : 244 – 253 .
Stanovnik , B. and Tisler , M. ( 1972 ) A new synthesis of pyrido[1,2-a]-1,3,5-triazines . Synthesis 6 : 308 – 309 .
Takei , K. , Sakakibara , H. and Sugiyama , T. ( 2001 ) Identifi cation of genes encoding adenylate isopentenyltransferase, a cytokinin biosynthesis enzyme, in Arabidopsis thaliana . J. Biol. Chem. 276 : 26405 – 26410 .
Werner , T. , Motyka , V. , Laucou , V. , Smets , R. , Van Onckelen H. and Schmülling , T. ( 2003 ) Cytokinin-defi cient transgenic Arabidopsis plants show multiple developmental alterations indicating opposite functions of cytokinins in the regulation of shoot and root meristem activity . Plant Cell 15 : 2532 – 2550 .
Werner , T. , Motyka , V. , Strnad , M. and Schmülling , T. ( 2001 ) Regulation of plant growth by cytokinin . Proc. Natl Acad. Sci. USA 98 : 10487 – 10492 .
Yamada , H. , Suzuki , T. , Terada , K. , Takei , K. , Ishikawa , K. , Miwa , K. , et al . ( 2001 ) The Arabidopsis AHK4 histidine kinase is a cytokinin-binding receptor that transduces cytokinin signals across the membrane . Plant Cell Physiol. 42 : 1017 – 1023 .
Yoshida , S. , Forno , D.A. , Cock , J.H. and Gomez , K.A. ( 1972 ) Laboratory Manual for Physiological Studies of Rice , 2nd edn. pp. 53 – 57 . The International Rice Research Institute , Philippines .
2059
Cytokinin antagonists without agonist activity
Plant Cell Physiol. 51(12): 2047–2059 (2010) doi:10.1093/pcp/pcq163 © The Author 2010.
Dow
nloaded from https://academ
ic.oup.com/pcp/article/51/12/2047/1944222 by guest on 05 July 2022





























