Embed Size (px)
Citation preview

05.04.2013
1
Populationsgenetik
Zuchtgeschichte

05.04.2013
2
Ideale Population
• unendliche Populationsgröße
• Zufallspaarung (Panmixie)
• keine Mutationen
• keine Selektion
• keine Migration
Für die Häufigkeiten von Allelen und Genotypen gelten einfache mathematische Zusammenhänge.
Bei zwei Allelen je Genort sind drei Genotypen an einem Genort möglich:
A1 A1 A1 A2 A2 A2
Relative Genotypenhäufigkeit (Genotypenfrequenz):
Relative Häufigkeit eines Genotyps in der betrachteten Population
Genotyp Ni
Anzahl Tiere N= 0 - 1
p (A1 A1) + p (A1 A2 ) + p (A2 A2 ) = 1

05.04.2013
3
HARDY – WEINBERG - Regel
Eine Population befindet sich im genotypischen Gleichgewicht, wenn die beobachteten mit den erwarteten Genotypenfrequenzen übereinstimmen, die aus dem Binom der Allelfrequenzen berechnet werden.
(p + q)2 = p2 + 2 pq + q2
Bei Panmixie bleiben die Gen- und Genotypenfre-quenzen in einer unendlich großen Population über die Generationen hinweg konstant.
Genfrequenz für Ai = p
Genfrequenz für ai = qp + q = 1
p ( Ai ) q ( ai )
p (A) p2 (AA) pq (Aa)
q (a) pq (Aa) q2 (aa)
Häufigkeit = p2 + 2 p q + q2
p (A1 A1 ) = p2 = p ( A1 ) x p ( A1 ) = p x p = p2
p (A1 A2 ) = 2 pq = p ( A1 – Eiz.) x p (A2 – Sperm.) + p (A2 – Eiz.) x p (A1 – Sperm.)
pq + pq = 1pq
p (A2 A2 ) = q2 = q ( A2 ) x q ( A2 ) = q x q = q2
05.04.2013
4
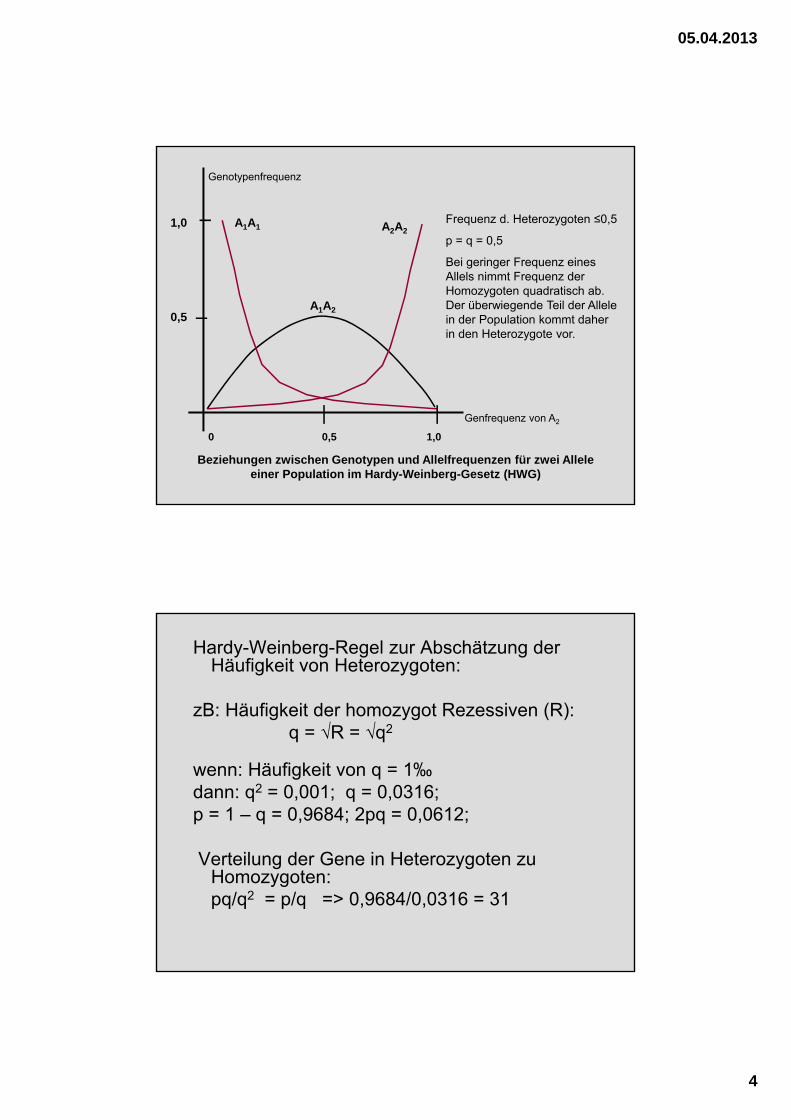
Beziehungen zwischen Genotypen und Allelfrequenzen für zwei Allele einer Population im Hardy-Weinberg-Gesetz (HWG)
A1A1 A2A2
A1A2
1,0
0,5
0 0,5 1,0
Frequenz d. Heterozygoten ≤0,5
p = q = 0,5
Bei geringer Frequenz eines Allels nimmt Frequenz der Homozygoten quadratisch ab. Der überwiegende Teil der Allele in der Population kommt daher in den Heterozygote vor.
Genfrequenz von A2
Genotypenfrequenz
Hardy-Weinberg-Regel zur Abschätzung der Häufigkeit von Heterozygoten:
zB: Häufigkeit der homozygot Rezessiven (R):q = √R = √q2
wenn: Häufigkeit von q = 1‰dann: q2 = 0,001; q = 0,0316;p = 1 – q = 0,9684; 2pq = 0,0612;
Verteilung der Gene in Heterozygoten zu Homozygoten:pq/q2 = p/q => 0,9684/0,0316 = 31

05.04.2013
5
Sonderfall : „geschlechtschromosomal gebundene Gene“
• Allelfrequenzen der O Nachkommen können direkt ausgewählt werden (auch bei vollst. Dominanz) Kennküken
• Mütter haben gleiche Allelfrequenz außer bei Mutation, Selektion, Migration
• Allelfrequenzen d. O Nachkommen = x der Eltern
• Allelfrequenzen der gesamten Nachkommenschaft hat zu 2/3 die der Mutter, 1/3 die des Vaters (y-Chromosom hat keine Allele)
Bedeutung der HARDY-WEINBERG-Regel für die praktische Tierzucht
Ohne Störung“ ändert sich die vorhandene Genfrequenz nicht
Eine Generation reicht aus, um nach Panmixie das gestörte Gleichgewicht wieder herzustellen (für 1 Gen gültig !)
Bei Panmixie ändern sich Allel- und Genfrequenzen in aufeinanderfolgenden Generationen nicht
Voraussetzung: Elterngeneration ist im Gleichgewicht
gültig auch bei Reinzucht
Prüfung der Frage, ob Zufallspaarung in der Population stattfand. Wirkung der Selektion

05.04.2013
6
Änderung der Genfrequenz:
Mutation Spontane Änderung der Erbsubstanz
MigrationEin- und Auswanderung von Genotypen (Zuchttierzukauf, Zuchttierverkauf)
Selektion Natürliche Selektion: Bevorzugung von Genen, die für die Arterhaltung förderlich sind
Künstliche Selektion: Zuchttierabhängige Bevorzugung einzelner Eltern
Genetische DriftZufällige Veränderungen in kleinen Populationen
Mutation
Mutationsrate: Wahrscheinlichkeit mit der 1 Gen in einer Generation mutiert
10- 4 bis 10 - 8
nur geringe Veränderung der Genfrequenz
Bei einmaliger Mutation besteht eine sehr geringe Wahrscheinlichkeit, dass das mutierte Gen in der Population erhalten bleibt.
Mutation beim rezessiven Gen: Verlustrisiko in der nächsten Generation ist sehr hoch
Mutiertes Gen kann Selektionsvorteil bzw. –nachteil für die Tiere hervorrufen, die es besitzen (Anlagen- /Merkmalsträger)

05.04.2013
7
Mutation
Bei 1xiger Mutation geringe Wahrscheinlichkeit, dass das mutierte Gen in der Population erhalten bleibt.
Hohes Verlustrisiko in den nächsten Generationen !
oder
Mutiertes Gen ruft Selektionsvorteil hervor für die Tiere, die es besitzen.
Wahrscheinlichkeit mit der 1 Gen in einer Generation mutiert:
= Mutationsrate
Bei Mensch und höheren Tieren: 10-4 bis 10 -8
nur sehr geringe Veränderung der Genfrequenz
Formen der Mutation
Genommutation: Euploidie Aneuploidie
ganzzahlige nichtganzzahlige
Veränderung des Chromosomensatzes
Chromosomenmutation: Veränderung der Struktur der Chromosomen
Deletion...........Fehlen von Teilstücken
Duplikation.......Teilstücke verdoppelt
Inversion..........Drehung einzelner Abschnitte (1800)
Translokation...Verlagerung von Abschnitten auf
nicht homologe Chromosomen
Genmutation: Veränderung innerhalb eines Gens
Bedeutung für die Tierzüchtung
große und kleine Mutationen Haustierwerdung !

05.04.2013
8
Späte somatische Mutation
Frühe somatische Mutation
Beginn
Zellpopulation
Endzustand
Zeitpunkt des Entstehens der
somatischen Mutation bestimmt den Anteil
der mutierten Zellen im betroffenen Gewebe
Etablierte und spontane Chromosomenmutationen
Strukturelle1 Numerische2+
Mutationsformen
Geschlechts-chromosomen
Autosome Chromosomen
- Geschlechts-organentwicklung wird beeinflusst
- Fortpfl.-störungen
-Phänotypveränderung
- Syndrome
-Fortpfl.störung
- Fitnessreduktion
- Kopplungsbrüche
- Fruchtb.-probleme
- Embryonalsterblichkeit
- Letalität
- Anatomische Missbild.
+
a) Etablierte Mutation: neben dem Normalkaryotyp auftretende andere Formen, z.B. Zentromerfusionen, die in best.Frquenzen in Populationen vorkommen
b) Spontane Mutation: neue Mutation (de novo)
1) Strukturelle Mutationen: Translokation, Inversion, Duplikation Deletion, Fusionen
2) Numerische Mutationen: Monosomien, trisomien, Triploidien, Polyploidien
05.04.2013
9
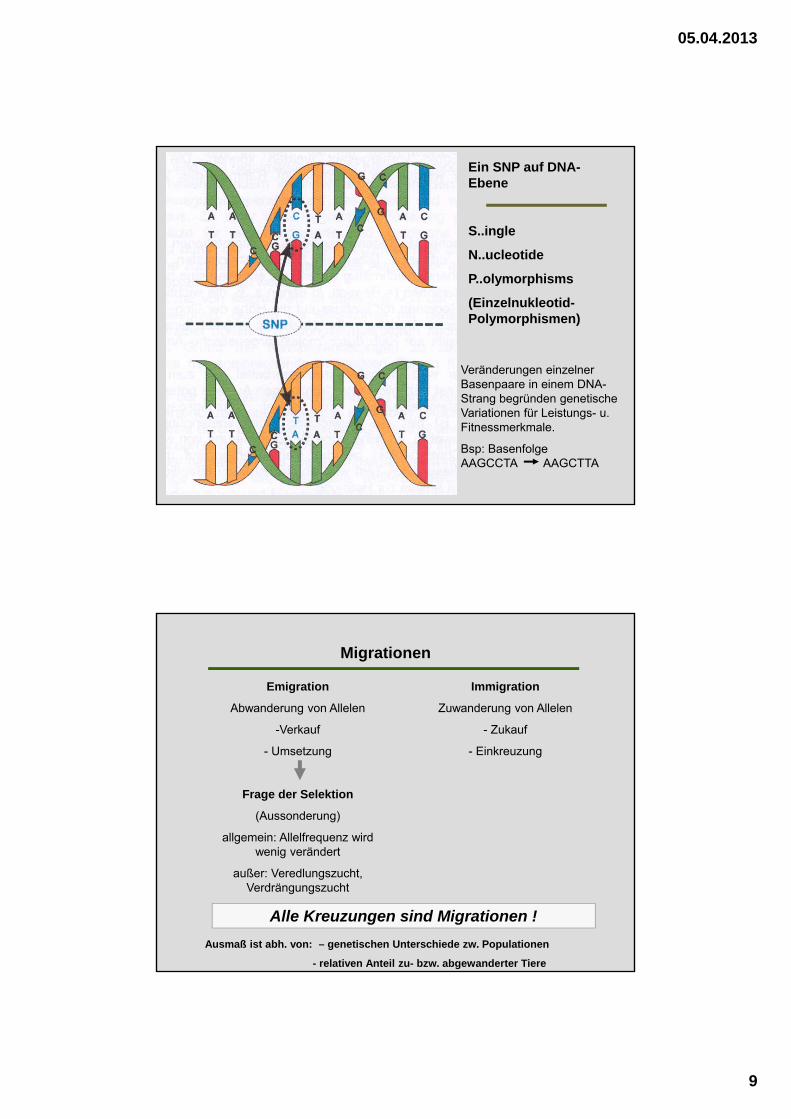
Ein SNP auf DNA-Ebene
S..ingle
N..ucleotide
P..olymorphisms
(Einzelnukleotid-Polymorphismen)
Veränderungen einzelner Basenpaare in einem DNA-Strang begründen genetische Variationen für Leistungs- u. Fitnessmerkmale.
Bsp: Basenfolge AAGCCTA AAGCTTA
Migrationen
Emigration
Abwanderung von Allelen
-Verkauf
- Umsetzung
Frage der Selektion
(Aussonderung)
allgemein: Allelfrequenz wird wenig verändert
außer: Veredlungszucht, Verdrängungszucht
Immigration
Zuwanderung von Allelen
- Zukauf
- Einkreuzung
Alle Kreuzungen sind Migrationen !
Ausmaß ist abh. von: – genetischen Unterschiede zw. Populationen
- relativen Anteil zu- bzw. abgewanderter Tiere
05.04.2013
10
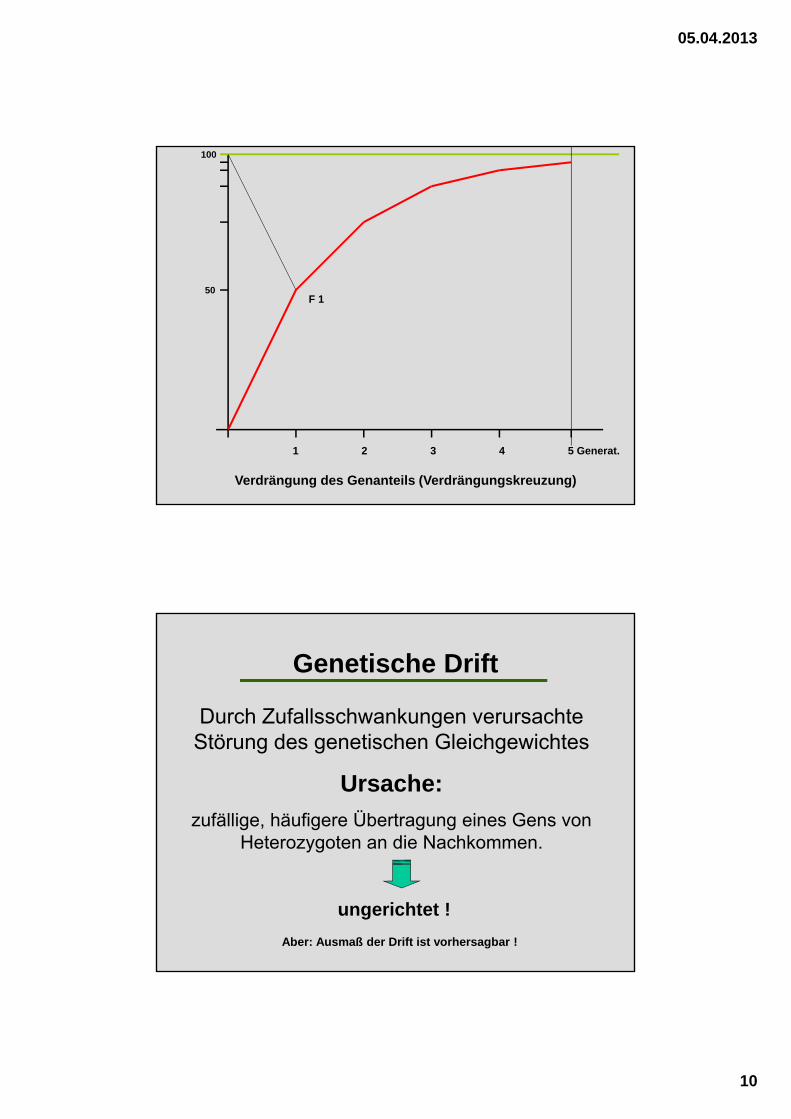
Verdrängung des Genanteils (Verdrängungskreuzung)
1 2 3 4 5 Generat.
100
50F 1
Genetische Drift
Durch Zufallsschwankungen verursachte Störung des genetischen Gleichgewichtes
Ursache:
zufällige, häufigere Übertragung eines Gens von Heterozygoten an die Nachkommen.
ungerichtet !
Aber: Ausmaß der Drift ist vorhersagbar !

05.04.2013
11
Begründung der Ursache: Heterozygotieanteil
Von beiden Genen an einem Locus wird nur ein Gen an Nachkommen weitergegeben.
Zufällige Weitergabe:
Homozygotie
ohne Einfluss
Heterozygotie
Bei zufälliger Bevorzugung des einen Gens Veränderung des genetischen Gleichgewichts
In einer großen Population ist die zufällige Bevorzugung eines Gens nicht möglich, weil das Gen entsprechend seines Erwartungswertes weitergegeben wird.
Große Population entspricht etwa einer Population !
Genfrequenzen werden infolge der Genetischen Drift zu Zufallsvariablen
Standardabweichung:
p =pq
2N
p, q : Genfrequenz
N : Populationsumfang
P = q = 0,5
Driftbedingte Genfrequenzschwankungen sind allgemein reversibel.
d.h. sie „driften zurück“
Zufallsschwankungen können Elimination des einen und Fixation des anderen Gens bewirken. In kleinen Population ist die Wahrscheinlichkeit der Fixation hoch. Langfristig vermehrte Homozygotie Inzucht !
Führt zu genetischen Differrenzierung zwischen Populationen !

05.04.2013
12
Einflussgrößen auf das Ausmaß der genetischen Drift
- Populationsumfang
- Anteil Heterozygoten
Homozygotie- /Inzuchtsteigerung je Generation durch zufällige genetische Drift ist von der Populationsgröße abhängig.
1
2NF = N = Anzahl der Elterntiere
Inzuchtkoeffizient nach t-Generationen mit Drift:
Ft = 1 – (1 – F)
Effektive Populationsgröße
Anzahl von Individuen in einer Idealpopulation, die der Anzahl Individuen in einer realen Population mit unterschiedlichem
Geschlechterverhältnis entspricht.
1 1 1
Ne 4Nm 4Nw
= +
Nm Anzahl männlicher Individuen
Nw Anzahl weiblicher Individuen
Ne effektive Populationsgröße
bei Nm : Nw = 1 Ne = N
in praxi (KB): Ne < N Beispiele
Nm x Nw
Nm + Nw
Ne = 4

05.04.2013
13
Beispiel: Rinderzucht (KB)
1 Bulle
400 Kühe
1 1 1
Ne 4 1600= + = 0,250625
1
2NeF = = 0,128
Inzuchtsteigerung je Generation
Ne = 3,99
Beispiel SchweinezuchtN = 50 Tiere
Beispiel Nm Nw Ne F%* F 10**
1 2 48 7,7 6,5 48,9
2 5 45 18,0 2,8 24,7
3 10 40 36,0 1,4 13,2
4 20 30 48,0 1,04 9,9
5 25 25 50,0 1,0 9,6
* F =1
2Ne** F 10 = 1 - (1 - F)10

05.04.2013
14
Einfluss des Geschlechtsverhältnisses auf die effektive Populationsgröße
Nm 1 10 1 2 1 2 1 2
Nw 1 10 10 10 100 100 1000 1000
Ne 2 20 3,64 6,67 3,96 7,84 4,00 7,98
Nm x Nw
Nm + Nw
Ne = 41 1 1
Ne 4Nm 4Nw
= +
Qualitative Genetik

05.04.2013
15
Qualitative Merkmale:
• nur von einem bzw. wenigen Genpaaren beeinflusst
• kaum bzw. nicht von Umwelteffekten beeinflussbar
• alternatives Auftreten
• vererben sich nach den MENDELschen Gesetzen
Quantitative Merkmale:
• von mehreren Genpaaren beeiflusst
• relativ umweltabhängig
• fließende Übergänge
• durch Maß und Zahl erfassbar
Erbfehler
Erbfehler sind erblich bedingte, morphologische oder physiologische unerwünschte Abweichungen von der Norm, die die Lebensfähigkeit, den Zuchtwert, den Gebrauchswert der Tiere beeinträchtigen.
Erbfehler sind meist die Folge von Mutationen an einzelnen Genorten. Sie werden sichtbar, wenn die allelen Genorte beider Chromosomen rezessiv besetzt sind.
Erbfehler werden meist mono- oder difaktoriell vererbt. Häufig tritt unvollständige Penetranz auf.

05.04.2013
16
Erbfehler
Letalfaktoren:
100 %iger Tod vor Erreichen der Geschlechtsreife
Semiletalfaktoren:
Tod bei 50 – 99 % der Tiere vor Erreichen der Geschlechtsreife
Subvitalfaktoren:
Tod bei 0 – 49 % der Tiere vor Erreichen der Geschlechtsreife
Populationsgenetik
Erbfehler (Erbdefekte)
Erbmängel
geringe
Erbkrankheiten
(Erbleiden)
schwere
Rasse-,
Zuchtfehler
Nicht krankhafte
Abweichungen vom
Rassestandard
(zB. rot bei SB)
gesundheitl. Störungen

05.04.2013
17
Häufigkeit von Erbfehlern beim Schwein
Anteil d. Frequenz Frequenz (%)
HodensackbruchNabelbruchNachhandlähmungZwitterbildungAfterverschlussDickbeinigkeitBinneberKarpfengebissHirnbruchWasserkopfKrüppelohrStülpzitzen
46,0412,4410,609,228,765,534,611,380,460,460,46
0,730,200,170,150,140,090,070,020,010,010,01
bis 0,05100,0
Erbfehler beim Rind:
• Bovine Leukozyten Adhäsion Defizienz (BLAD)
•Bovine progressive degenerative Myeloencephalopathie (Weaver)
•Uridin - 5‘ - Monophosphat - Synthetase (DUMPS)

05.04.2013
18
Letalfehler des Rindes und deren Erbgänge
ymbol Name Erbgang Symbol Name Erbgang
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22
Achondroplasie (Zwergwuchs) Epitheliogenesis imperfecta (Hautdef.) Achondroplasie Hypotrichosis congenita (Haarlosigkeit) Acroteriasis congenita (Fehlen v. Gliedm.) Mumifikation der Frucht Nachhandparalyse (Stehunvermögen) Muskelkontraktur Kieferankylose (Gelenkverknöcherung) Wirbelsäulenverkürzung Ljutikows Letalfaktor Angeborene Wassersucht Allg. Ankylose (Gelenkverknöcherung) Mißbildung d. Molaren (Kieferverkürzung) Achondroplastische Kleingliedrigkeit Atresia ani (Afterverschluss) Missbildung d. Beine (verdreht, verknöchert) Hernea cerebralis (Hirnbruch) Brachygnathia inferior (Unterkieferverkürzg.)Agnathie (Kieferlosigkeit) Geschl.-gebundener Letalfaktor Beiderseitger Nasenverschluss
Dom.Rez rez. rez. rez. rez. rez. rez. rez. rez. rez. rez. rez. rez. rez. rez. rez. rez. rez. rez. rez. rez.geschdom.
A 23 A 24 A 25 A 26 A 27 A 28 A 29 A 30 A 31 A 32 A 33 A 34 A 35 A 36 A 37 A 38 A 39 A 40 A 41 A 42 A 43
Fehlen v. Hintergliedm. (Kriechkälber) Hydrocephalus (Wasserkopf) Angebor. Spasmen u. Ataxie Verlängerte Tragezeit 1 Verlängerte Tragezeit 2 Strichweise Haarlosigkeit (weibl.) Achondroplasie Angeborene Porphyrinurie Dysfunkt. d. Thyroidea (Kopfverkürz Ichthyosis congenita (Hyperkeratose) Anadontia (Haar- u. Zahnlosigkeit) Aplasie d. Hypophysenvorderlappens Erbl. Muskelkontr. (Beinverdrehg) Erbl. Lähmg. Hinterbeine mit Blindheit Reduz. Wirbelzahl verl. Dornforts. Polyzythämie Atresia ilei (Darmverschluss) Probatozephalie (Hammelkopf) Spaltbildung d. Wirbelsäule Abort in Graviditätsmitte Parakeratose (Störg. Zn-Stoffw.)
rez. rez. Rez Rez. rez.. dom rez. rez. ? rez. rez. rez. rez. rez. rez. rez. rez. ? rez. rez. rez.
rez.
Zum Heterozygotietest autosomal rezessiv deter-minierter Merkmale sind grundsätzlich geeignet:
• Anpaarung an weibliche Merkmalsträger (nur bei Lebensfähigkeit derselben)
• Anpaarung an bekannte Heterozygote
• Anpaarung an Töchter bekannter Heterozygoter
• Inzuchttest (Vater - Töchterpaarungen, Vollgeschwisterpaarungen
• Anpaarung an Probanden der allgemeinen Population („automatische Prüfung“ oder„Praxismethode“
praktisch nicht durch-führbar

05.04.2013
19
Häufigkeit des Auftretens von Erbfehlern
Abhängig von: - Mutationsrate
- Selektionsdruck gegen Erbfehler
Mutationsrate = Selektionskoeffizient xHäufigkeit unerwünschter Gene
= s x q2
q =
s Bei s = 1: q =
q0
1 + nq0
qn =
Häufigkeit in der n-ten = Generation
Häufigkeit in der Anfangsgeneration
1 + nq0
Berechnung der Verminderung der Merkmalsträger in nachfolgenden Generationen
Bedingung: genereller Zuchtausschluss von Merkmalsträgern
(rezessiv letal + semiletal)
05.04.2013
20
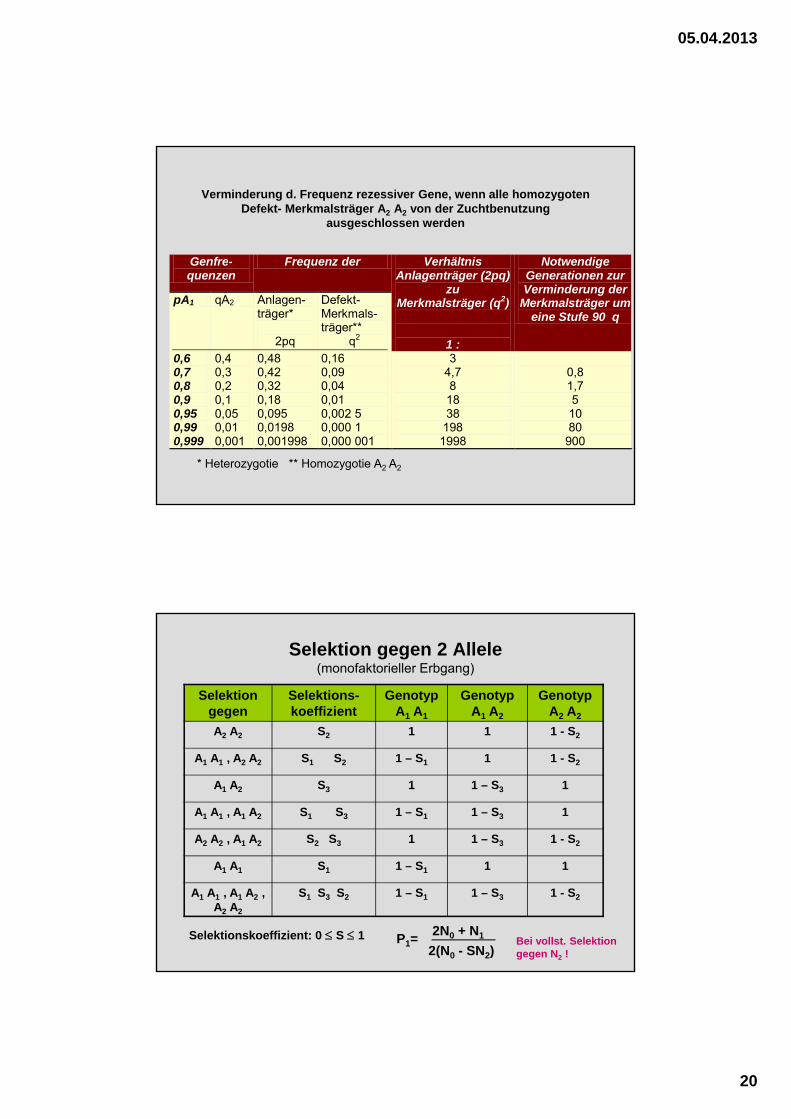
Verminderung d. Frequenz rezessiver Gene, wenn alle homozygoten Defekt- Merkmalsträger A2 A2 von der Zuchtbenutzung
ausgeschlossen werden
Genfre-quenzen
Frequenz der
pA1 qA2 Anlagen-träger*
2pq
Defekt-Merkmals-träger**
q2
Verhältnis Anlagenträger (2pq)
zu Merkmalsträger (q2)
1 :
Notwendige Generationen zur Verminderung der
Merkmalsträger um eine Stufe 90 q
0,6 0,7 0,8 0,9 0,95 0,99 0,999
0,4 0,3 0,2 0,1 0,05 0,01 0,001
0,48 0,42 0,32 0,18 0,095 0,0198 0,001998
0,16 0,09 0,04 0,01 0,002 5 0,000 1 0,000 001
3 4,7 8 18 38
198 1998
0,8 1,7 5 10 80
900 * Heterozygotie ** Homozygotie A2 A2
Selektion gegen 2 Allele (monofaktorieller Erbgang)
Selektion gegen
Selektions-koeffizient
Genotyp A1 A1
Genotyp A1 A2
Genotyp A2 A2
A2 A2 S2 1 1 1 - S2
A1 A1 , A2 A2 S1 S2 1 – S1 1 1 - S2
A1 A2 S3 1 1 – S3 1
A1 A1 , A1 A2 S1 S3 1 – S1 1 – S3 1
A2 A2 , A1 A2 S2 S3 1 1 – S3 1 - S2
A1 A1 S1 1 – S1 1 1
A1 A1 , A1 A2 , A2 A2
S1 S3 S2 1 – S1 1 – S3 1 - S2
Selektionskoeffizient: 0 S 1 P1=2N0 + N1
2(N0 - SN2)Bei vollst. Selektion gegen N2 !

05.04.2013
21
Populationsgenetik
• Ursachen von Letalfaktoren / Erbfehler:– Rein genetisch
– Wechselwirkung Gene mit Umwelt
– Rein exogen (Phänokopie)
• Anzahl von Letalfaktoren:
Ca. 2000 bei Menschen
Ca. 230 bei Rindern
Ca. 70 bei Schweinen
Ca. 50 beim Pferd
Quantitative Genetik

05.04.2013
22
Heritabilität
Heredity = Erblichkeit................h2
Verhältnis d. genotypischen zur phänotypischen Varianz
Anteil der genetisch bedingten Varianz an der Gesamtvarianz
S2
S2h2 = =
S2
S2 + S2
g g
p g u
h2 im engeren Sinne: im Zähler wird nur die additiv genetische Varianz verwendet
h2 im weiteren Sinne:im Zähler ist die gesamte genetische Varianz aufgeführt. S (additiv) + S (dominant) + S (epistatisch)
0 < h2 < 1
Erblichkeit:biologischer Begriff der genetischen Verankerung von Merkmalen
Erblichkeitsgrad:genetisch statistischer Begriff in welchem Maße phänotypische Varianzen auf genetische Ursachen zurückzuführen sind
Bedeutung von h2: Züchten dient der Verbesserung des genetisch begründetes Leistungspotentials
Wichtig !
Phänotypische Rangfolge der Tiere = genotypische Rangfolge
Das Maß der Übereinstimmung zwischen der phänotypischen und genotypischen Rangfolge ist h2.
= Maß der Zuverlässigkeit der möglichen Selektionsmaßnahmen
direkter Einfluss auf den Selektionserfolg

05.04.2013
23
Erhöhung der Heritabilität (h2) durch:
Maßnahmen:
• Leistungsprüfung unter einheitlichen Bedingungen
• Verringerung von Meßfehlern
• Relativierung auf einen Vergleichsmaßstab
• Verwendung von Vollgeschwistermittelwerten statt Einzelleistungen
•Verwendung mehrerer korrespondierender Leistungen
Verringerung der Umweltvarianz !
h2 - Werte für einzelne Merkmalsgruppen
h2 Merkmale
Schwach< 0,25
Fruchtbarkeit, Fitness, Resistenz, absoluteMilchmengenleistung, Persistenz
Mittelstark0,25 - 0,60
Masteigenschaften < Schlachteigenschaften, relativeMilchmenge u. Milchinhaltsstoffe, durchschnittlicheEinzeleimasse, Wollertrag, Reinwollgehalt
Stark> 0,60
Konformationsmaße, Knochenmaße, Strichlänge

05.04.2013
24
Wiederholbarkeit
Umweltbedingte Varianz
systematische Varianz
wirken auf mehrere Individuen gleichzeitig
kann statistisch eliminiert werden
zufällige Varianz
nicht fassbar
Verringerung durch mehrmaliges Messen
Varianz zw. Individuen (VB)
Varianz zw. Individuen (VB) + Varianz zw. Wiederholungen (VW)
W = (VB)
(VW)
Heritabilitätsschätzung
Zwei Methoden:
Schätzung der genetischen Varianz auf der Grundlage der Eltern -Nachkommen - Ähnlichkeit
Berechnung über den erreichten Zuchtfortschritt

05.04.2013
25
Eltern - Nachkommen - Analyse
Grundlage: Zurückfallen der Nachkommen (-leistung) zum Mittelwert der Eltern
Regression zum Elterndurchschnitt
Verwandtschaftsgrade: Elter Nachkomme..........0,50
Vollgeschwister..........................0,50
Halbgeschwister.........................0,25
Eineiige Zwillinge........................1,00
Verwandte sind einander ähnlicher als Nichtverwandte
Annahme: phänotyp. Ähnlichkeit zwischen Verwandten beruht allein auf abstammungsidentische Gene
h2 wird aus dem Grad der Ähnlichkeit zwischen Verwandten geschätzt
Heritabilitätsschätzung
Zwei Methoden:
Schätzung der genetischen Varianz auf der Grundlage de Eltern - Nachkommen
- Ähnlichkeit
Berechnung über den erreichten Zuchtfortschritt

05.04.2013
26
Berechnung der phänotypischen Ähnlichkeit zwischen Geschwistern
- Kovarianz zwischen phänotypischen Leistungen*
* Kovarianz: Beschreibung der Abhängigkeit zwischen 2 Merkmalen - Wertepaare x, y
SXY = (xi - y) ( yi - y)1
n - 1
n
i-1
Schätzung der berechneten Kovarianzen: VHG = 0,25 VA
Eltern-NK = 0,5 VA
VG = 0,5 VA + 0,25 VD
VA = additive Varianz
VD = dominanzbedingte Varianz
Eltern - Nachkommen - RegressionEN 1 VA 1
2E 2 VP 2
bE,N = = = h2
Berechnung der EN-Kovarianzen
EN = (xE - xE)(xN - xN)
n
n
i=1
Selektion
Natürliche Selektion
Auswahl der Individuen nach dem Grad der
Fitness
Künstliche Selektion
Ziel:
gerichtete Selektion stabilisierende Selektion disruptive Selektion
Objekt:
Individualselektion Familienselektion direkte Selektion indirekte Selektion
Auswahl von erwünschten, für die Weiterzucht geeigneten Individuen oder Familien und die Ausmerzung von nicht
geeigneten Individuen oder Familien aus einer Population.

05.04.2013
27
Beispiel für Allelfrequenzänderung durch Selektion
Herde mit 100 Tieren enthält 4 Albinos (A 2, A2) davon sind 52 Tiere heterozygot (A1, A2) und 44 Tiere homozygot (A1, A1)
Ziel ist die vollständige Selektion der Albinos.
Formel für Berechnung der Allelfrequenz
2N0 + N1
2(N - sN2)P =
N0 Anzahl beobachteter Individuen bei A1 A1 N1 bei A1 A2 N2 bei A2 A2 N Gesamtzahl an Individuen s = Selektion
Ausgangssituation: p = = 0,72 x 44 + 52
2 x 100
Nach Selektion der 4 Albinos: p = = = 0,7292 x 44 + 52
2 (100 - 1 x 4)
140 A1 - Allele
192 A - Allele
Allelfrequenzänderung von p = - p = 0,729 -0,700 = 0,029
d.h.: Allel A2 vermindert sich um 2,9 %
Allel A1 erhöht sich um 2,9 % !
korrelierte Selektion Simultanselektion S. nach einem Merkmal S. nach mehreren Merkmalen S. nach unabhängigen S.-grenzen S. nach voneinander abhängigen S.-grenzen
Selektionsverfahren :
Selektion
Natürliche Selektion Künstliche Selektion
• Verschiebung des Populationsmittels
• Änderung der Genotypenfrequenz
• Einschränkung der Varianz
• Verringerung der Anpassungsfähigkeit

05.04.2013
28
Künstliche Selektion
Gezielte Auswahl von Individuen einer Population zur Weiterzucht.
Ziel:
Auswahl der leistungsmäßig besten Individuen mithilfe einer möglichst großen Selektionsdifferenz zur Erzielung eines möglichst hohen Selektionserfolges in der nächsten Generation.
Wirkung:
Änderung der Allelfrequenz in der nächsten Generation.
Selektionsdifferenz
Mittlere Leistungsüberlegenheit der selektierten Eltern gegenüber dem Mittel der gesamten Elternpopulation.
Grundlage der SD sind phänotypische Merkmale im Rückschluss auf genotypische Werte.
= ausgedrückt in Einheiten der phänotypischen Standardabweichungen der betreffenden Merkmale
Die so standardisierte SD = Selektionsintensität i
05.04.2013
29
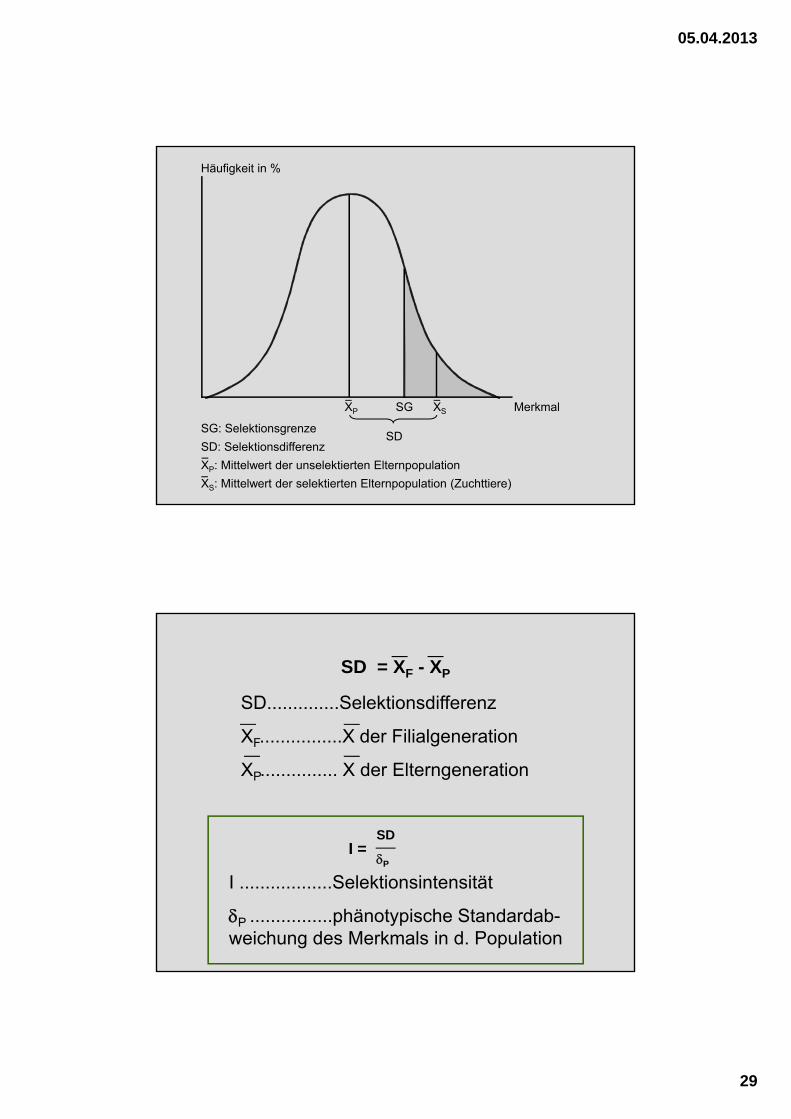
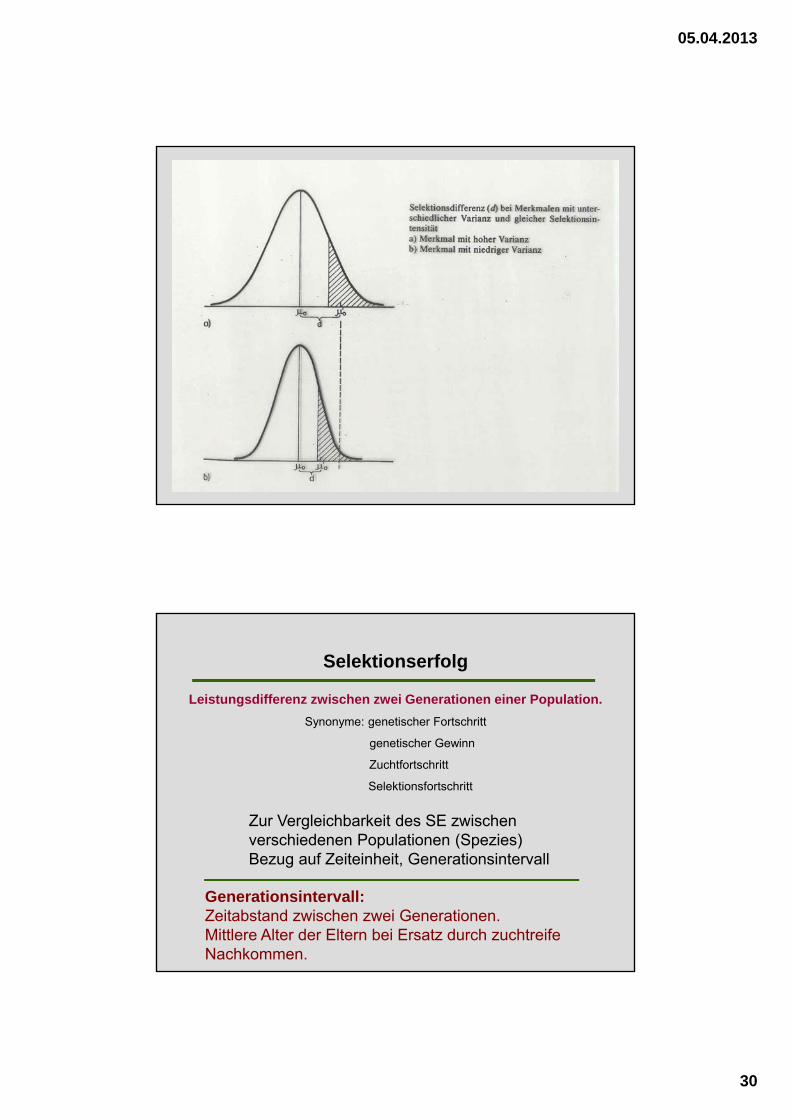
SG
SD
Merkmal
Häufigkeit in %
SG: Selektionsgrenze
SD: Selektionsdifferenz
XP XS
XP: Mittelwert der unselektierten Elternpopulation
XS: Mittelwert der selektierten Elternpopulation (Zuchttiere)
SD = XF - XP
SD..............Selektionsdifferenz
XF................X der Filialgeneration
XP............... X der Elterngeneration
I = SD
P
I ..................Selektionsintensität
P ................phänotypische Standardab-weichung des Merkmals in d. Population
05.04.2013
30
Selektionserfolg
Leistungsdifferenz zwischen zwei Generationen einer Population.
Synonyme: genetischer Fortschritt
genetischer Gewinn
Zuchtfortschritt
Selektionsfortschritt
Zur Vergleichbarkeit des SE zwischen verschiedenen Populationen (Spezies) Bezug auf Zeiteinheit, Generationsintervall
Generationsintervall: Zeitabstand zwischen zwei Generationen. Mittlere Alter der Eltern bei Ersatz durch zuchtreife Nachkommen.

05.04.2013
31
Überlegenheit der Eltern lässt sich nicht 100 %ig auf Nachkommen übertragen !
Heritabilität
(0 h2 1)
h2 =Sg
2
Sg2 + Sp
2
Sg2 .... genet. Varianz
Sp2 .... phänotyp. Varianz
SE = h2 x SD
h2 x SD
LSE =
S ........ Regression vom Mittel der selektierten Eltern auf das Mittel ihrer Nachkommen
Heritabilität (0 h2 1)
h2 x SD
LSE =
=
SD Nachkommen je Zeiteinheit / Generation
Nutzungsdauer
L Pubertätsalter
Zuchtreife
Anzahl Nachkommen für die Zucht
05.04.2013
32
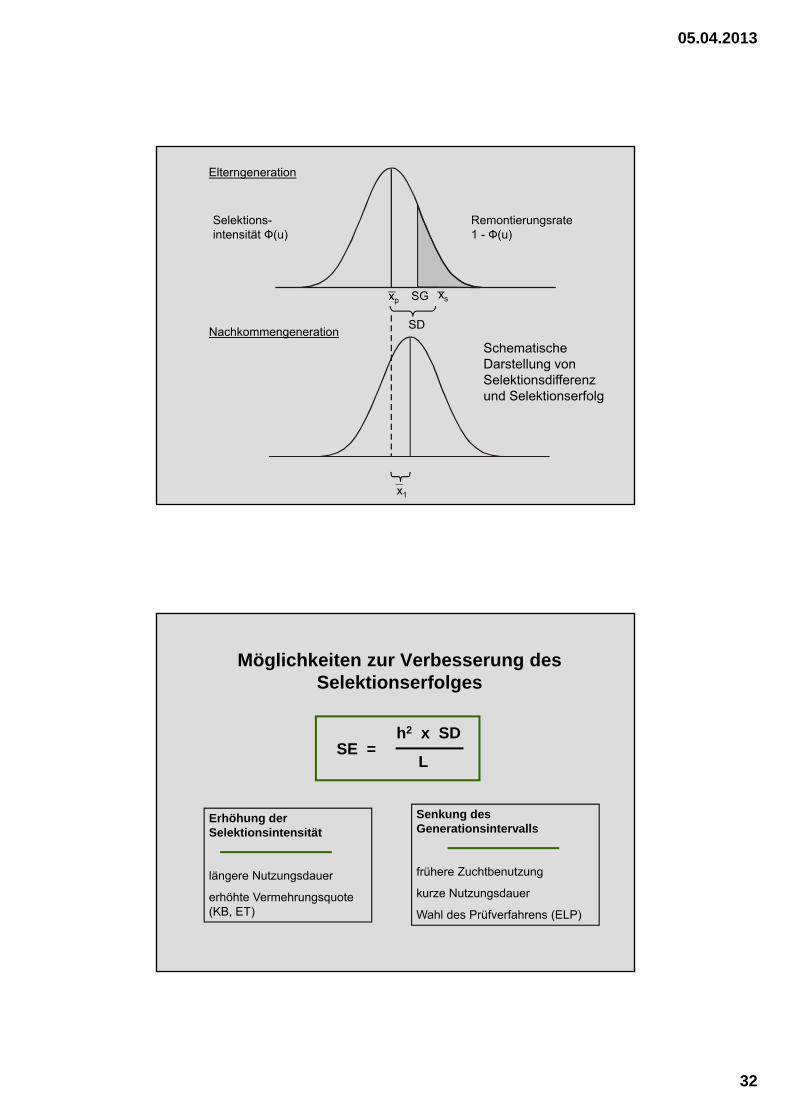
Elterngeneration
Nachkommengeneration
Selektions-intensität Ф(u)
Remontierungsrate1 - Ф(u)
SD
SGxpxs
x1
Schematische Darstellung von Selektionsdifferenz und Selektionserfolg
Möglichkeiten zur Verbesserung des Selektionserfolges
h2 x SD
LSE =
Erhöhung der Selektionsintensität
längere Nutzungsdauer
erhöhte Vermehrungsquote (KB, ET)
Senkung des Generationsintervalls
frühere Zuchtbenutzung
kurze Nutzungsdauer
Wahl des Prüfverfahrens (ELP)

05.04.2013
33
SE
SD
h2
I
Zahl d. Selektionsmerkmale
Merkmalsvariation
Selektionsintensität
Genetisch bedingte Variation
Umweltbedingte Variation
Genauigkeit der Zuchtwertschätzung
Nutzungsdauer
Frühreife
Zeitpunkt der Selektion
Genetische Korrelation zw. Merkmalen Volkswirtschaftliche Zielstellung Zuchtziel
Kreuzung / Reinzucht Rassenkonsolidierung
Bedarf an Nachzucht Zahl der Nachkommen
Populationsgröße Kreuzung / Reinzucht
Ernährung Haltung Nutzung
Genetische Variation Leistungsniveau Schätzverfahren / InformationsquellenGesundheit Leistungsselektion
Genetische Veranlagung Ernährung
Genauigkeit der Zuchtwertschätzung Züchtungsaufwand
Populationsgröße genetische Ausgangssituation
Selektionsdauer
Genetisch nutzbare Variabilität
Heritabilität h2
Selektions-erfolg
SESelektionsdifferenz (SD) Selektionsintensität (SI)
Merkmale Remontierungs-prozentsatz
Gen. Korrelation zw. Merkmalen Wirtschaftliche
Ziele
Anpaarungs-verhältnis Aufzuchtergebnis
Generationsintervall
Nutzungsdauer Vermehrungsrate
Legereife Pubertät
Zuchtreife
Bestimmungskomponenten des Selektionserfolges
05.04.2013
34
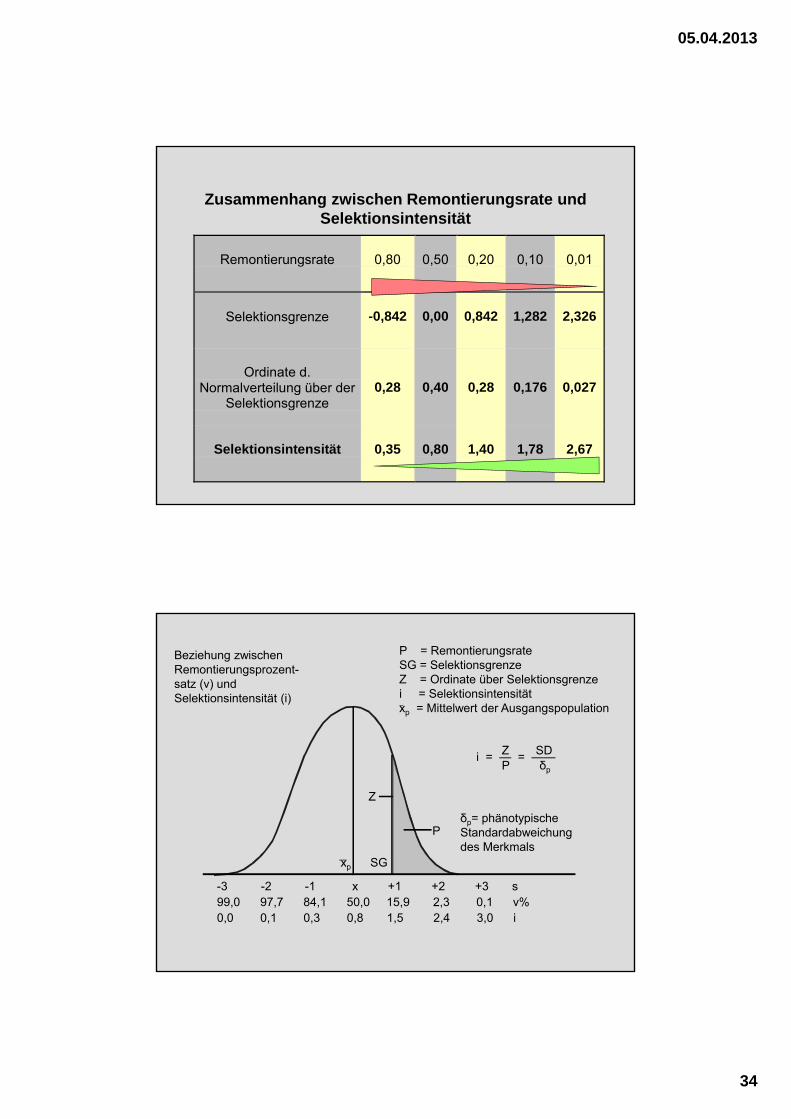
Zusammenhang zwischen Remontierungsrate und Selektionsintensität
Remontierungsrate 0,80 0,50 0,20 0,10 0,01
Selektionsgrenze -0,842 0,00 0,842 1,282 2,326
Ordinate d.Normalverteilung über der
Selektionsgrenze0,28 0,40 0,28 0,176 0,027
Selektionsintensität 0,35 0,80 1,40 1,78 2,67
δp= phänotypische Standardabweichung des Merkmals
Z
SGxp
i = — = ——Z SDP δp
-3 -2 -1 x +1 +2 +3 s99,0 97,7 84,1 50,0 15,9 2,3 0,1 v%0,0 0,1 0,3 0,8 1,5 2,4 3,0 i
P = RemontierungsrateSG = SelektionsgrenzeZ = Ordinate über Selektionsgrenzei = Selektionsintensitätxp = Mittelwert der Ausgangspopulation
P
Beziehung zwischen Remontierungsprozent-satz (v) und Selektionsintensität (i)
05.04.2013
35
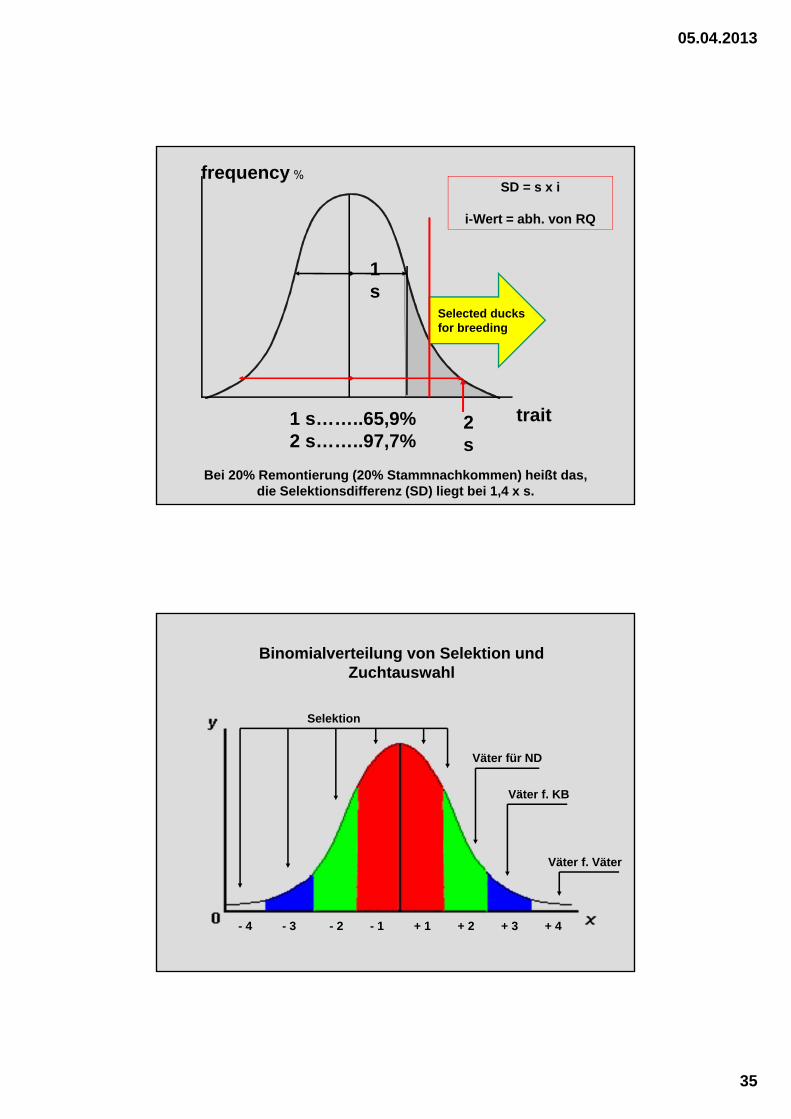
trait
frequency %
1s
2s
1 s……..65,9%2 s……..97,7%
Selected ducks for breeding
Bei 20% Remontierung (20% Stammnachkommen) heißt das, die Selektionsdifferenz (SD) liegt bei 1,4 x s.
SD = s x i
i-Wert = abh. von RQ
Selektion
Väter für ND
Väter f. KB
Väter f. Väter
- 4 - 3 - 2 - 1 + 1 + 2 + 3 + 4
Binomialverteilung von Selektion und Zuchtauswahl

05.04.2013
36
Wechselwirkung zwischen Selektionsintensität und Generationsintervall (Bsp. Schweinezucht)
Nutzungsdauer der Sau 1 Wurf 5 Würfe
2 weibl. Ferkel /Wurf 2 10
Remontierung 50 % 10 %
Generationsintervall 1 Jahr 3 Jahre
SE + 25 %
Kriterien für Leistungsmerkmale zwecks Selektionswürdigkeit innerhalb der Reinzucht
• Wirtschaftliche Bedeutung
• Höhe der Heritabilität (h2)
• genetische Korrelation zu anderen Merkmalen
• nutzbare additive genetische Varianz
• Genauigkeit der Merkmalserfassung (Aufwand - Nutzen - Relation)

05.04.2013
37
Einflussfaktoren auf den Erfolg der Selektion
• Intensität der Selektion
• Anzahl der gewählten Leistungsmerkmale
• Genauigkeit der Zuchtwert- bzw. Erbwertschätzung
• Anteil der additiv genetisch bedingten Variation des entsprechenden Merkmals
SE =1
n
0
20
40
60
80
100
120
1 2 3 4 5 6 7 8 9 10
SE
Der relative Selektionserfolg (SE) je Merkmal reduziert sich mit zunehmender Anzahl an Merkmalen, die gleichzeitig berücksichtigt werden !
Anzahl Merkmale Selektionserfolg / Merkmal (%)
1 100,0
2 70,7
3 57,7
5 44,7
8 35,4
10 31,6
15 25,8

05.04.2013
38
Tandemselektion
Merkmale werden nacheinander züchterisch berücksichtigt !
1. Merkmal bis Ziel erreicht
2. Merkmal bis Ziel erreicht
Je Merkmal ist eine unterschiedliche Anzahl an Generationen erforderlich !
Nachteil:
Rangierung der Merkmale nach ökon. Bedeutung ist kaum möglich
Nichtberücksichtigung von Merkmalen führt zum Leistungsabfall zum Mittelwert hin !
Vorteil:
Selektion erfolgt nur nach einem Merkmal. Das gestattet einen hohen Selektionserfolg.
Selektion nach unabhängigen Selektionsgrenzen
Festlegung eines Mindestniveaus für jedes Merkmal, d.h. genereller Ausschluss von Individuen, die in einem Merkmal die Mindestanforderungen nicht erfüllen.
Praktische Durchführung:
• Beginn mit einem der n Merkmale
• aus der positiv selektierten Teilpopulation wird nach dem 2. Merkmal selektiert
• Fortgesetzte Wiederholung dieser Vorgehensweise bis zum n-ten Merkmal
1 Merkmal.........R
2 Merkmale.... R
....
n Merkmale.... Rn
Vorteil:
leichte Anwendbarkeit
Nachteil der Tandemselektion wirkt nicht
Nachteil:
evtl. Ausschluss von sehr guten Zuchttieren, die nur in einem Merkmal die Selektionsgrenze nur knapp nicht erreichen.

05.04.2013
39
Gesamtpopulation
Teilpopulation 1
Teilpopulation 2
Zuchtpopulation
Selektion nach unabhängigen Selektionsgrenzen
1. Merkmal
2. Merkmal
3. Merkmal
Remontierungsquoten
für 1 Merkmal..............R
für 2 Merkmale........ R
für n Merkmale........ Rn
Bsp. 25 % Zuchttiere stehen zur Selektion zur Verfügung. Die Auswahl erfolgt nach 3 Merkmalen.
0,25 = 0,63
63 % werden nach dem 1. Merkmal, 63 % nach dem 2. Merkmal, 63 % nach dem 3 Merkmal selektiert.
14
Selektion nach voneinander abhängigen Selektionsgrenzen
(Indexselektion)
Nachteile der Selektion nach unabhängigen Selektionsgrenzen wirken nicht.
Berücksichtigung mehrerer Merkmale. Merkmale sind voneinander abhängig.
Nachteil:
Zuchtfortschritt je Einzelmerkmal ist mit wachsender Anzahl an Merkmalen zunehmend begrenzt.

05.04.2013
40
Selektionsindex
I = (b1 x1 ) + (b2 x2) +..........+ (bn xn)
I..................Index (dimensionslos)
b.................Regressionskoeffizient des Merkmals xn1, der abhängig ist
von der ökonomischen Bedeutung (Grenznutzen), der
Heritabilität, der verwandtschaftlichen Beziehung zw.
Proband und Informant sowie von den genetischen u.
phänotypischen Korrelationen zwischen den im Index
enthaltenen Merkmalen
x1...xn..........individuelle phänotypische Abweichung des Merkmals
1..n des Probanden vom Mittelwert des Vergleichsmaßst.
T = ai x Gi
n
I = 1
ai .......ökon. Gewicht
Gi......Teilzuchtwert
T = 7,5GAF + 0,06GTZ – 48 GFV + 7,5 GWT + 1,5 GFB
Universalzuchtwert für Fleischschweine
T = 0,06GTZ – 48GFV + 7,5GWT + 3GFB
TVL = 0,06GTZ – 48GFV + 9,75GWT + 1,5GFB
TML= 7,5GAF + 0,06GTZ – 48 GFV + 7,5 GWT + 1,5 GFB
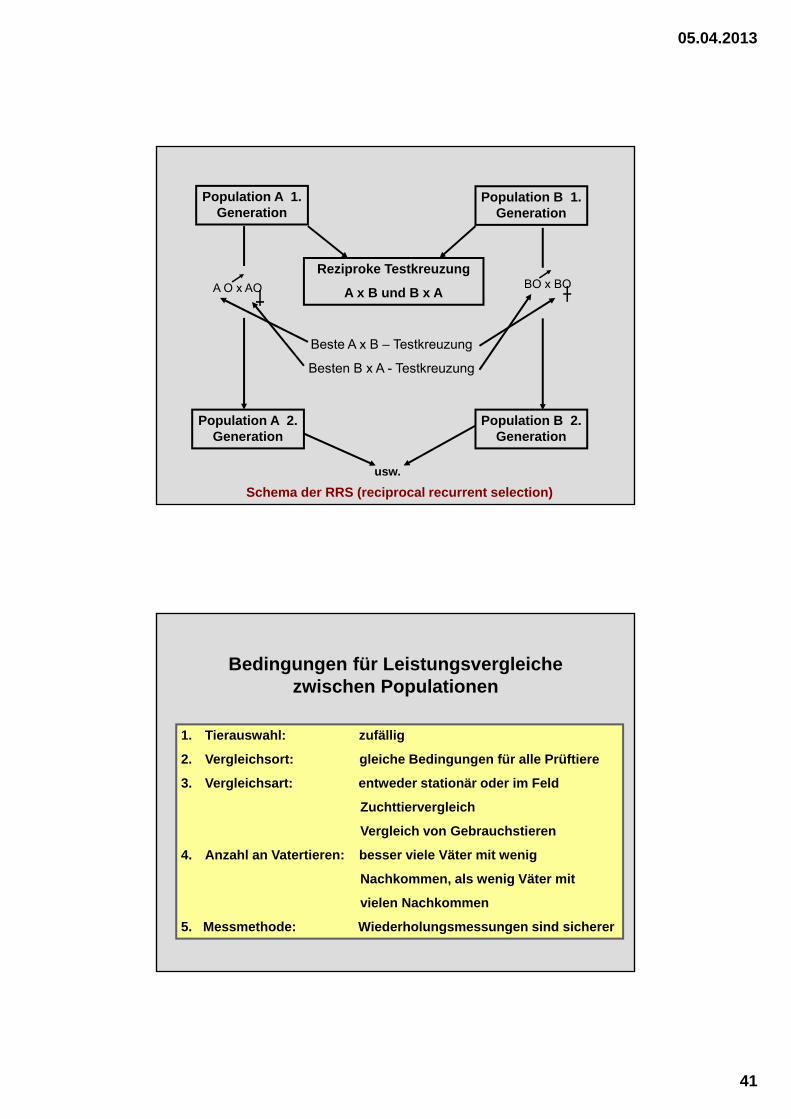
05.04.2013
41
Population A 1. Generation
Population B 1. Generation
Reziproke Testkreuzung
A x B und B x AA O x AO BO x BO
Beste A x B – Testkreuzung
Besten B x A - Testkreuzung
Population A 2. Generation
Population B 2. Generation
Schema der RRS (reciprocal recurrent selection)
usw.
Bedingungen für Leistungsvergleiche zwischen Populationen
1. Tierauswahl: zufällig
2. Vergleichsort: gleiche Bedingungen für alle Prüftiere
3. Vergleichsart: entweder stationär oder im Feld
Zuchttiervergleich
Vergleich von Gebrauchstieren
4. Anzahl an Vatertieren: besser viele Väter mit wenig
Nachkommen, als wenig Väter mit
vielen Nachkommen
5. Messmethode: Wiederholungsmessungen sind sicherer

05.04.2013
42
Verwandtschaft und Inzucht
Zwei Individuen sind verwandt, wenn sie gleiche Gene besitzen.
Paarung verwandter Individuen = Inzucht
Paarung nicht verwandter Individuen = Fremdzucht
Inzucht Einengung der genetischen Variabilität
Fremdzucht Erweiterung d. genetischen Variabilität
Verengung
Erweiterung
des Erbgutes
Sel
bst
bef
ruch
tun
g
Vat
er x
To
chte
r B
rud
er x
Sch
wes
ter
Hal
bg
esch
wis
terp
aaru
ng
2 g
emei
nsa
me
Gro
ßel
tern
1 g
emei
nsa
mer
Gro
ßel
ter
Kei
ne
gem
ein
sam
e A
hn
en
bis
…G
ener
atio
nen
Ras
seg
leic
he
Tie
re
vers
chie
den
er H
erku
nft
Ras
sekr
euzu
ng
Art
kreu
zun
g
Pfe
rd x
Ese
l
Gat
tun
gsk
reu
zun
g
Sch
af x
Zie
ge
Inzucht
Zufällige Paarung
Fremdzucht
Blutauffrischung
Reinzucht Kreuzung

05.04.2013
43
Inzuchtkoeffizient bei fortgesetzter Inzucht
Inzucht-generation
Verpaarungsysteme
Vollgeschwister Eltern-Nachkommen
Halbgeschwister
1 0,250 0,250 0,125
2 0,375 0,375 0,219
3 0,500 0,438 0,305
4 0,595 0,469 0,381
5 0,672 0,484 0,449
20 0,986 0,500 0,904
Inzuchtgrad = Verwandtschaftskoeffizient
=
Wahrscheinlichkeit, dass zwei Gene am gleichen
Locus der Eltern identisch sind und sich im
Nachkommen vereinigt haben
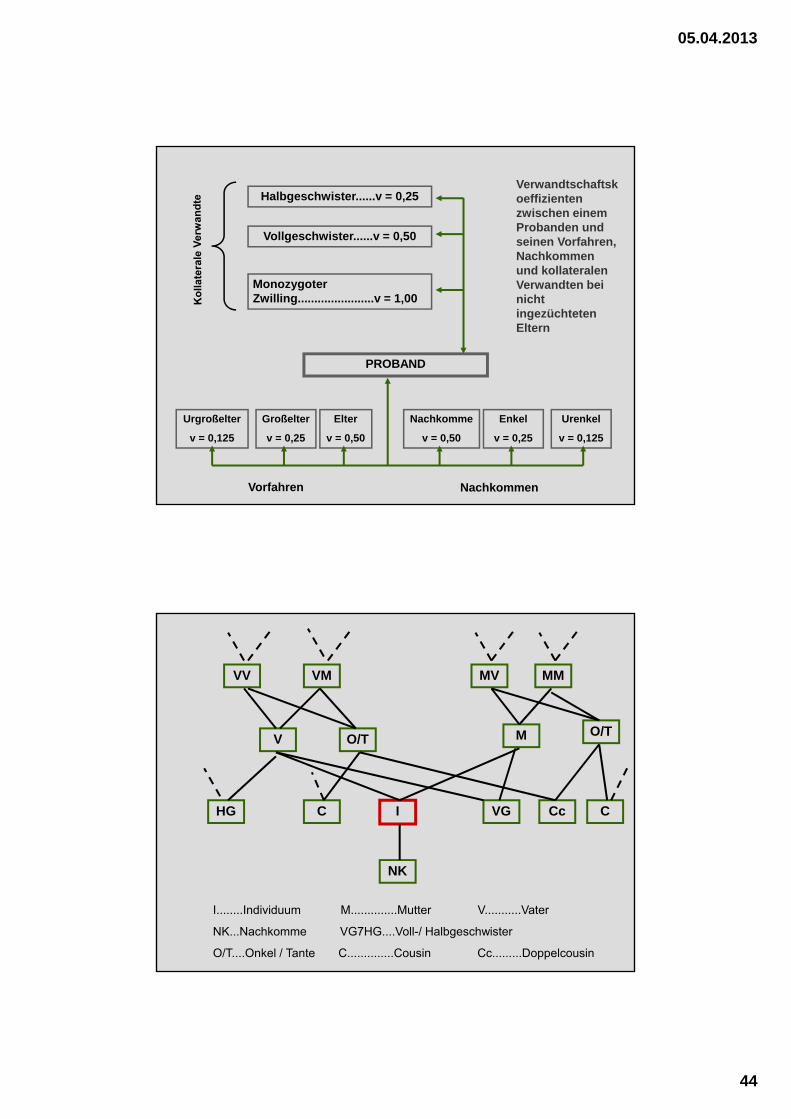
05.04.2013
44
Halbgeschwister......v = 0,25
Vollgeschwister......v = 0,50
Monozygoter Zwilling.......................v = 1,00
PROBAND
Urgroßelter
v = 0,125
Großelter
v = 0,25
Elter
v = 0,50
Nachkomme
v = 0,50
Enkel
v = 0,25
Urenkel
v = 0,125
Vorfahren Nachkommen
Verwandtschaftskoeffizienten zwischen einem Probanden und seinen Vorfahren, Nachkommen und kollateralen Verwandten bei nicht ingezüchteten Eltern
VV
CcVGHG
V
NK
I
O/T
VM
M O/T
MMMV
I........Individuum M..............Mutter V...........Vater
NK...Nachkomme VG7HG....Voll-/ Halbgeschwister
O/T....Onkel / Tante C..............Cousin Cc.........Doppelcousin
C C

05.04.2013
45
Wenn mehrere gemeinsame Ahnen vorkommen, dann summieren sich die Wahrscheinlichkeiten
für den Besitz gleicher Gene !
Einfache Ermittlung der additiv genetischen Verwandtschaft
z.B. Vollgeschwister: Pfad I – V – VG = 0,5 x 0,5 = 0,25
Pfad I – M – VG = 0,5 x 0,5 = 0,25
insgesamt: 0,50
Verwandtschaft zwischen I und Ahne = 0,5n
n .....Anzahl Generationen zw. I u. Ahne
Verwandtschaftskoeffizient:Wahrscheinlichkeit, dass zwei Gene am gleichen Locus identisch sind
Locus befindet sich an 2 verschiedenen Individuen Verwandtschaftsgrad
Locus befindet sich am gleichen Individuum Inzuchtgrad
Der additiv genetische Verwandtschaftsgrad (a)
=
doppelter Verwandtschaftskoeffizient
Wenn 2 Individuen die herkunftsgleiche Chromosomen besitzen, miteinander verpaart werden, können Nachkommen auftreten, die diese in sich vereinen und demzufolge homozygot (für diese Chromosomen) sind.
Inzuchtgrad (FX) entspricht der Wahrscheinlichkeit, dass zwei Gene am gleichen Locus der Eltern von X identisch sind und in X sich vereinigt haben !
Inzuchtgrad FX = halber Verwandtschaftsgrad zwischen den Eltern !

05.04.2013
46
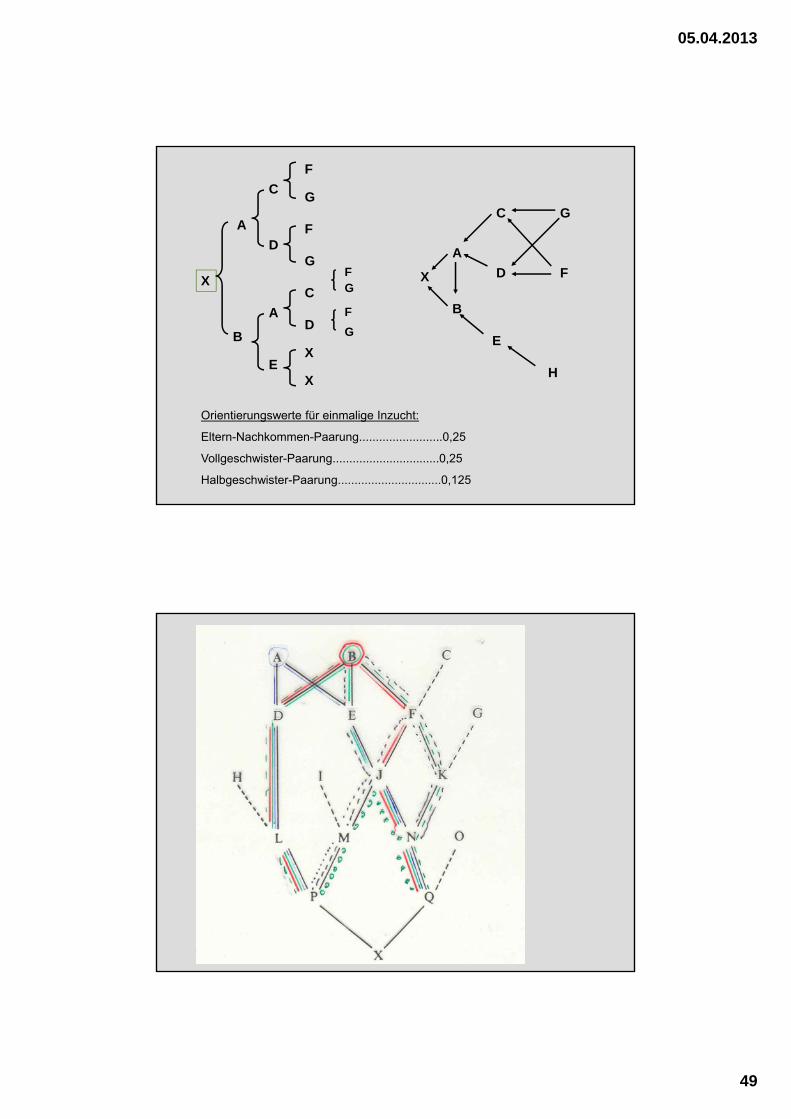
Berechnung des Inzuchtkoeffizienten nach der „Pfad-Methode“
• Unterteilung des Pedigree nach mütterlicher und väterlicher Linie
• Verbindung derjenigen Ahnen, die sowohl mütterlich als auch väterlich vorkommen, aber nur dann, wenn sie mütterlich und väterlich über mehrere Generationen verschiedene Nachkommen haben
• Sind gemeinsame Ahnen ingezüchtet ? Wenn ja, berechnet man den Inzuchtgrad separat und setzt ihn als F(x) ein.
• Rechenschema ausfüllen
• Regel 2 beachten:In der Kette „Vater – gemeinsamer Ahne – Mutter“ darf nicht zweimal der gleiche Ahne vorkommen !
X
B
C
D
E
D
F

05.04.2013
47
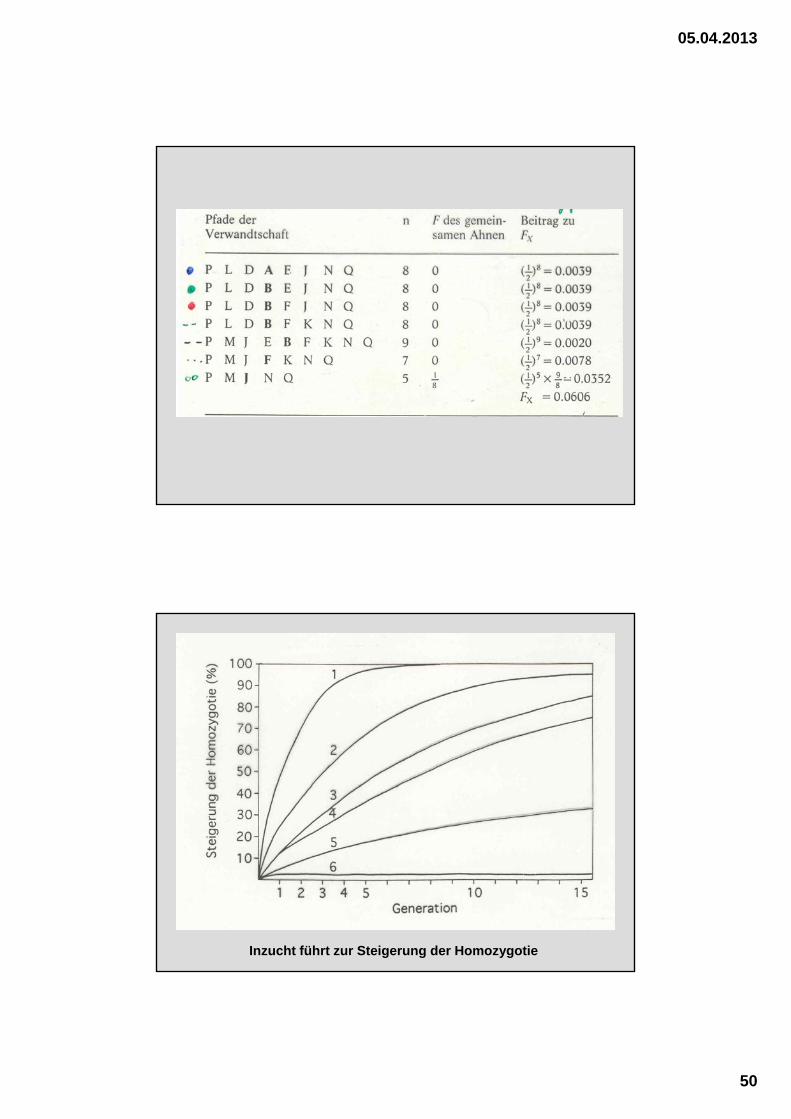
Schätzformel für die Berechnung des Inzuchtkoeffizienten
1. Gemeinsamer Ahne ist selbst nicht ingezüchtet
F(X) = 0,5 n1 + n2 + 1
n.........Anzahl an Generationen zw. Proband u. gemeinsamen Ahnen
1.......... väterliche Seite (d.h. vom Vater bis zum Ahnen)
2...........mütterliche Seite (d.h. von der Mutter bis zum Ahnen)
2. Gemeinsamer Ahne ist selbst ingezüchtet
F(X) = 0,5 n1 + n2 + 1 (1 + F(A))
F(A)............Inzuchtkoeffizient des Ahnen
Väterliche Abstammung Mütterliche Abstammung
H I H K - - - - G FH I - - G F H K
G F - - E - E F
E - C D
A B
X
Ist der Proband X auf mehrere (k) Ahnen ingezüchtet, dann berechnet man für jeden dieser Ahnen deren Inzuchtbeitrag nach der Formel
F(X) = 0,5 n1 + n2 + 1
und bildet die Summe dieser k Inzuchtbeiträge. Dabei beginnt man mit den Vorfahren, die den geringsten Zeitunterschied zum Probanden aufweisen.
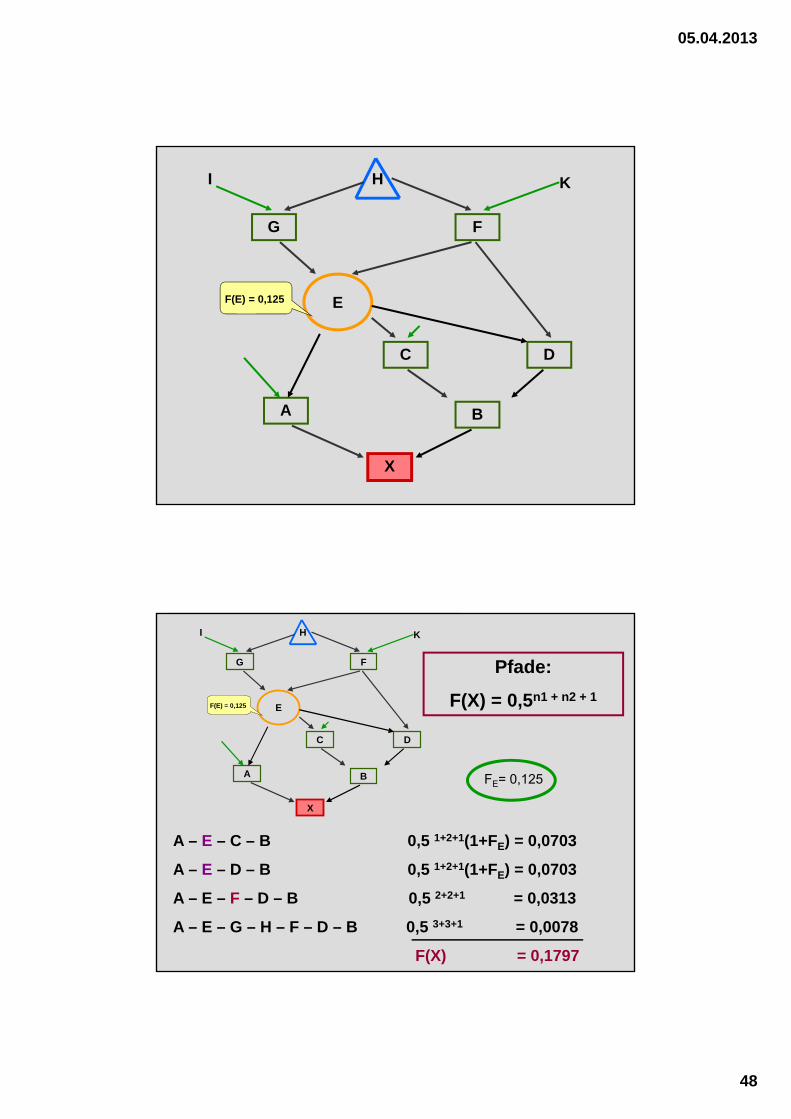
05.04.2013
48
X
A B
C D
E
F
H
G
I K
F(E) = 0,125
A – E – C – B 0,5 1+2+1(1+FE) = 0,0703
A – E – D – B 0,5 1+2+1(1+FE) = 0,0703
A – E – F – D – B 0,5 2+2+1 = 0,0313
A – E – G – H – F – D – B 0,5 3+3+1 = 0,0078
F(X) = 0,1797
X
A B
C D
E
F
H
G
I K
F(E) = 0,125
Pfade:
F(X) = 0,5n1 + n2 + 1
FE= 0,125
05.04.2013
49
X
A
C
B
D
A
E
G
F
C
F
G
D
X
X
FG
F
G
X
C
A
G
B
D F
E
H
Orientierungswerte für einmalige Inzucht:
Eltern-Nachkommen-Paarung.........................0,25
Vollgeschwister-Paarung................................0,25
Halbgeschwister-Paarung...............................0,125
05.04.2013
50
Inzucht führt zur Steigerung der Homozygotie

05.04.2013
51
Reguläre Inzuchtsysteme
Zur Züchtung von Inzuchtlinien werden oft bestimmte Verwandte über mehrer Generationen gepaart. Die Berechnung des erwarteten Inzuchtgrades erfolgt nach den sog. Rekursionsformeln (WRIGHT, 1921)
1. Selbsbefruchtung Ft = ½ (1 + Ft-1 )
2. Vollgeschwisterpaarung Ft = ¼ (1 + 2F t-1 +F t –2)
3. Halbgeschwisterpaarung Ft = 1/8 (1 + 6F t-1 + F t-2)
4. Doppelcousin Ft = 1/8 (1 + 4F t-1 + F t-2)
5. Cousin Ft = 1/8 (1 + 4F t-1 + 2F t-2 + F t-3)
6. Halbcousin
Inzuchtdepressionen bei 10%er Inzuchtsteigerung in der Population
• Milchleistung Rind.....................3 – 8 % (150 – 600 l)
• Wurfgröße Schwein...................ca. 5 % (0,5 Ferkel)
• Legeleistung Huhn....................ca. 4 % (10 Eier)
• Körpergewicht Broiler...............ca. 2 % (20 g)
05.04.2013
52
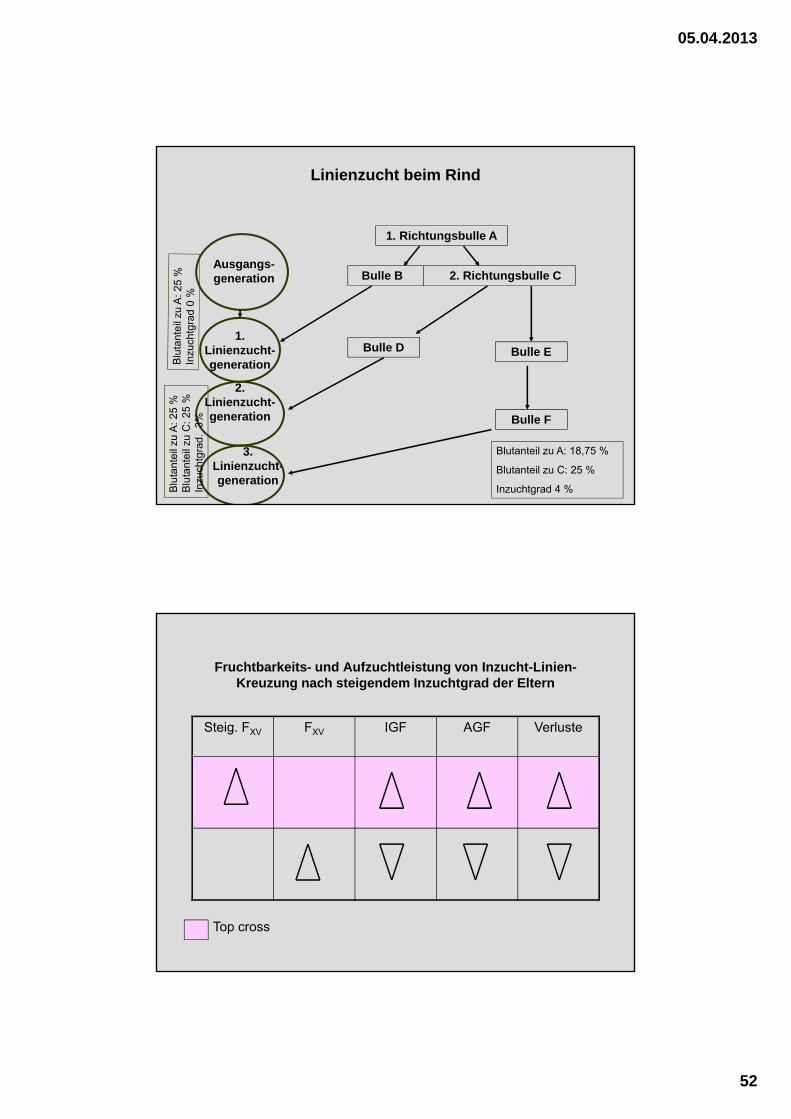
Linienzucht beim Rind
1. Richtungsbulle A
Bulle B 2. Richtungsbulle C
Bulle D
Bulle F
Bulle E
Ausgangs-generation
1. Linienzucht-generation
2. Linienzucht-generation
3. Linienzucht-generation
Blutanteil zu A: 18,75 %
Blutanteil zu C: 25 %
Inzuchtgrad 4 %Blu
tant
eil z
u A
: 25
%
Blu
tant
eil z
u C
: 25
%
Inzu
chtg
rad.
3%
Fruchtbarkeits- und Aufzuchtleistung von Inzucht-Linien-Kreuzung nach steigendem Inzuchtgrad der Eltern
Steig. FXV FXV IGF AGF Verluste
Top cross

05.04.2013
53
Gleichgewicht
Inzucht-depression
Heterosis-effekt
Inzucht Kreuzung
cis- eu- trans-
Heterosis
Zusammenhang zwischen Inzucht und Kreuzung sowie Inzuchtdepression und Heterosiseffekten
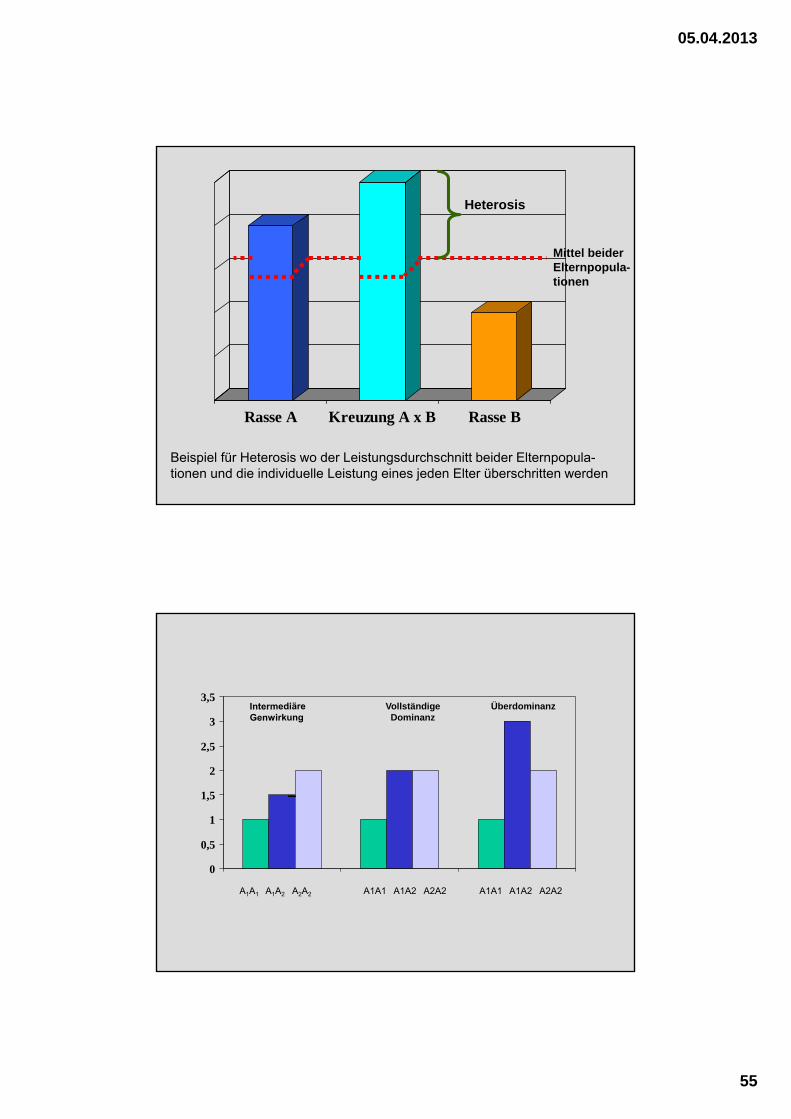
Heterosis
..wenn bei Kreuzungen die Leistungen der Nachkommen vom Mittel der beiden Eltern-populationen abweichen.
Diese Definition beinhaltet, dass Heterosis sowohl positiv als auch negativ sein kann.
H. ist die Aufhebung von Inzuchtdepression bei der Kreuzung von Inzuchtlinien im
engeren Sinne.

05.04.2013
54
Heterosis
....mittlere genetische Überlegenheit („hybrid vigour“)der Kreuzungs-nachkommen über den Durchschnitt der Elternpopulation bzw. über dem besten Elter.
Heterosiseffekt:
= die über das bei rein additiver Vererbung erwartete Niveau hinausgehende Leistungssteigerung
Heterosiseffekte sind bei der Kreuzung verwandt-schaftlich entfernter (genetisch distanzierter)
Populationen mit gleichen Merkmalen zu erwarten !
Ursachen für Heterosis
• Heterozygotie gilt als VoraussetzungHeterozygotie bewirkt physiologische Stimulierung
besseres Wachstumbessere Vitalität
• Dominanzeffekte verschiedener Genorte können zu positiven Wirkungen führen
• Überdominanzwirkungen zeigen besondere Wirkungen in Richtung Heterosis
• Bestimmte Formen der Epistasie können zu Heterosiswirkungen führen.
05.04.2013
55
Rasse A Kreuzung A x B Rasse B
Heterosis
Mittel beider Elternpopula-tionen
Beispiel für Heterosis wo der Leistungsdurchschnitt beider Elternpopula-tionen und die individuelle Leistung eines jeden Elter überschritten werden
0
0,5
1
1,5
2
2,5
3
3,5
A1A1 A1A2 A2A2 A1A1 A1A2 A2A2 A1A1 A1A2 A2A2
Vollständige Dominanz
Überdominanz
05.04.2013
56
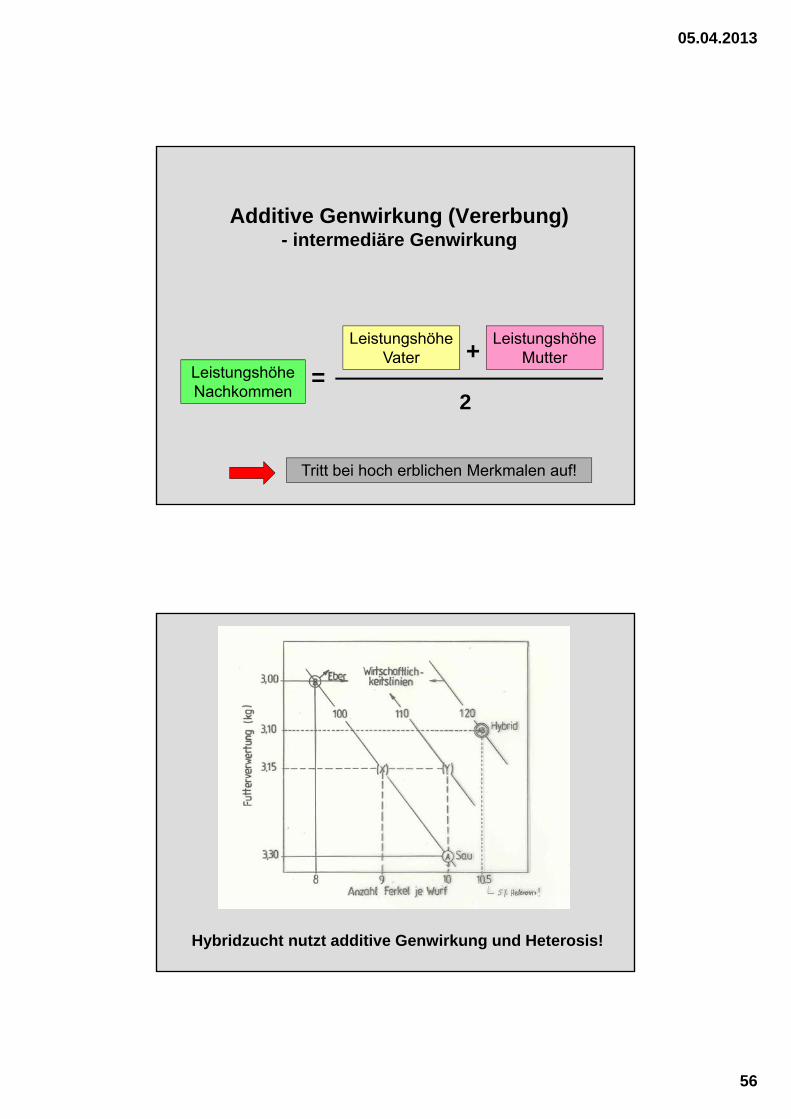
Additive Genwirkung (Vererbung)- intermediäre Genwirkung
Tritt bei hoch erblichen Merkmalen auf!
Leistungshöhe Nachkommen
Leistungshöhe Vater
Leistungshöhe Mutter
2
+=
Hybridzucht nutzt additive Genwirkung und Heterosis!
05.04.2013
57
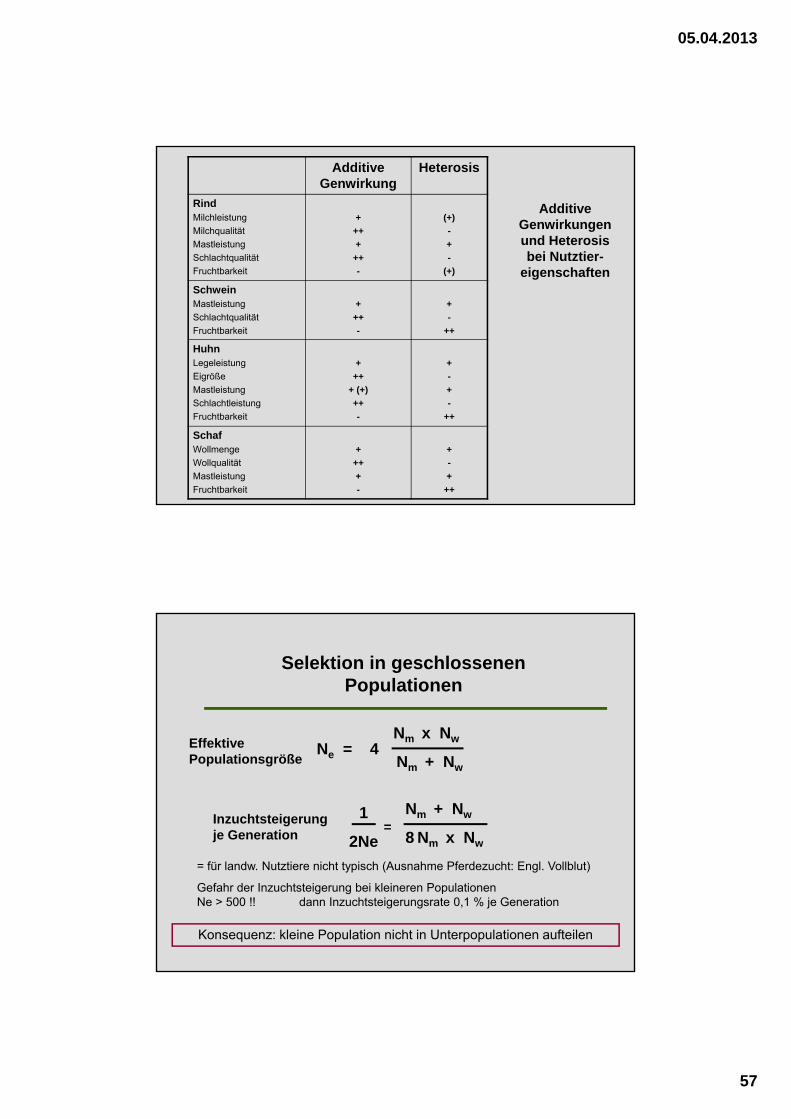
Additive Genwirkungen und Heterosis bei Nutztier-
eigenschaften
Additive Genwirkung
Heterosis
RindMilchleistung
Milchqualität
Mastleistung
Schlachtqualität
Fruchtbarkeit
+
++
+
++
-
(+)
-
+
-
(+)
SchweinMastleistung
Schlachtqualität
Fruchtbarkeit
+
++
-
+
-
++
HuhnLegeleistung
Eigröße
Mastleistung
Schlachtleistung
Fruchtbarkeit
+
++
+ (+)
++
-
+
-
+
-
++
SchafWollmenge
Wollqualität
Mastleistung
Fruchtbarkeit
+
++
+
-
+
-
+
++
Ne = 4Nm x Nw
Nm + Nw
Selektion in geschlossenen Populationen
Nm + Nw
8 Nm x Nw
1
2Ne=
Inzuchtsteigerung je Generation
Effektive Populationsgröße
= für landw. Nutztiere nicht typisch (Ausnahme Pferdezucht: Engl. Vollblut)
Gefahr der Inzuchtsteigerung bei kleineren Populationen Ne > 500 !! dann Inzuchtsteigerungsrate 0,1 % je Generation
Konsequenz: kleine Population nicht in Unterpopulationen aufteilen
05.04.2013
58
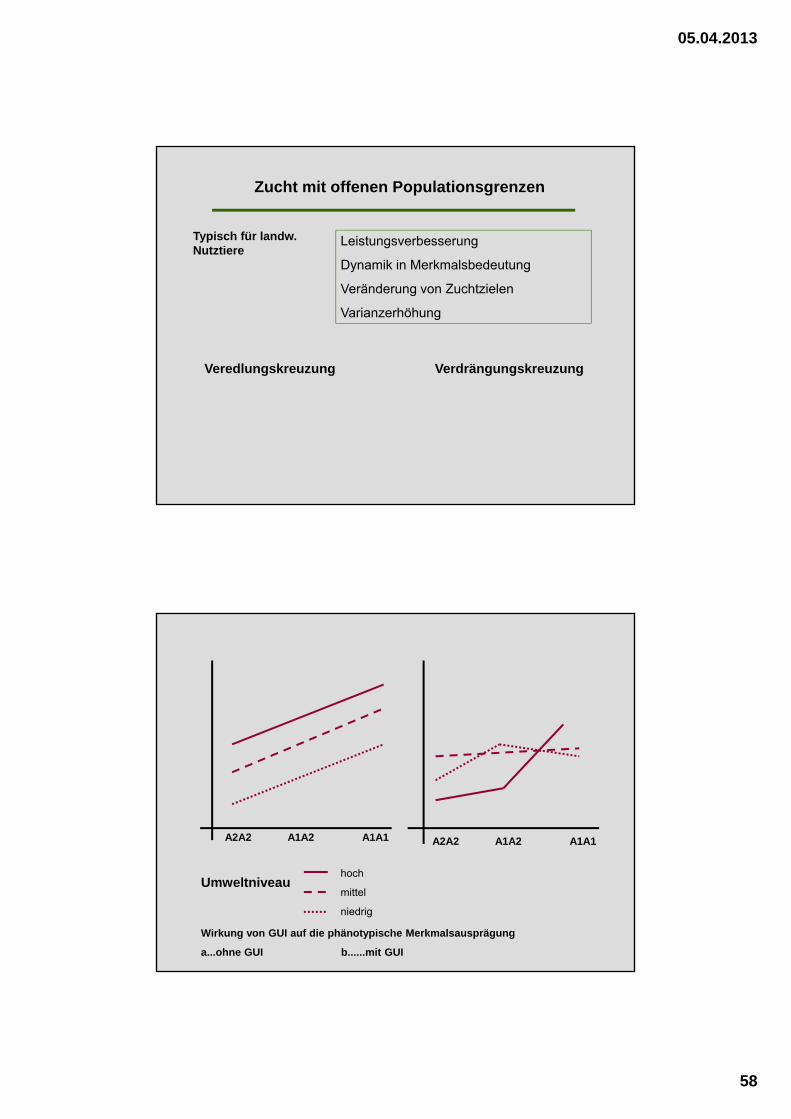
Zucht mit offenen Populationsgrenzen
Typisch für landw. Nutztiere
Leistungsverbesserung
Dynamik in Merkmalsbedeutung
Veränderung von Zuchtzielen
Varianzerhöhung
Veredlungskreuzung Verdrängungskreuzung
Wirkung von GUI auf die phänotypische Merkmalsausprägung
a...ohne GUI b......mit GUI
hoch
mittel
niedrig
Umweltniveau
A2A2 A1A2 A1A1 A2A2 A1A2 A1A1

05.04.2013
59
Genetik• Hauptentscheidungen mit größter Wirkung
auf künftige Gewinne
– Geographische Lage– Was ist der angepeilte Markt – kundenspezifische
Programme– Entscheidung zugunsten einer geschlossenen
Herde oder einer regulären Zuchtpyramide– Plan zur Herdengröße– Plan zum angestrebten Schlachtgewicht– Zusammenarbeit mit anderen an der
Produktionskette beteiligten Partner